


ISSN: 0973-7510
E-ISSN: 2581-690X
Candidatus Saccharibacteria (TM7) is an underexplored bacterial group within the human oral microbiome. These ultra-small, host-dependent organisms, despite their reduced genomes, significantly influence microbial balance and host immunity, exhibiting dual roles in oral health and disease and emerging as a key focus in microbiome research. This systematic review aimed to synthesize evidence on the prevalence, host interactions, genomic features, and ecological roles of TM7 in the oral cavity. A total of 13 peer-reviewed studies published between 2020 and 2025 were selected for detailed review based on predefined inclusion criteria, including clinical, laboratory, and metagenomic analyses focusing on TM7 detection, isolation, host specificity, and interactions with oral microbial communities. TM7 was found to be predominantly oral-adapted, exhibiting site-specific colonization with higher abundance in saliva. Coculture experiments demonstrated strict host dependence, particularly on Actinomyces, Schaalia, and Arachnia species, with strain-specific variations affecting host viability and microbial networks. TM7 genomes are highly reduced, with specialized adaptations including type IV pili and arginine deiminase pathways. Interactions with the immune system were context-dependent, with both immunomodulatory and pro-inflammatory outcomes observed. Detection methods significantly influenced TM7 prevalence and functional interpretation. Overall, TM7 represents a host-dependent, ecologically influential oral bacterium with genomic specialization, strain-level variability, and complex interactions within microbial communities. These findings highlight its potential roles in oral health and disease and emphasize the need for advanced multi-modal analytical approaches.
Candidatus Saccharibacteria, TM7, Oral Microbiome, Host Dependency, Metagenomics, Sequencing, Microbial Interactions
The human oral cavity has become a key model for studying microbial ecology and functionality due to its accessibility and clinical importance.1,2 Among the oral bacteria that are difficult to culture, Candidate Phyla Radiation (CPR) bacteria, particularly Saccharibacteria (TM7), have attracted considerable research attention. TM7 have been found as commensals across multiple body sites in healthy individuals, including the oral cavity, intestine, skin, and stomach.3 Some studies link members of the TM7 phylum with periodontal disease, whereas others suggest they may reduce bone loss and inflammation.1,2,4
The first observation of oral bacteria was made by Antonie van Leeuwenhoek in 1670, using his self-constructed microscope, marking the discovery of the oral microbiota.5,6 Subsequent research demonstrated that oral microorganisms significantly influence host health and that dysbiosis in the oral microbiome contributes to both oral and systemic diseases.7 The accessibility of oral microbiota allows direct observation of biofilm formation and microbial community assembly, making the oral cavity a powerful system for studying complex microbial ecosystems.8 Research indicates over 700 bacterial species inhabit the oral cavity, primarily from a few dozen genera across seven phyla: Actinomycetota (formerly Actinobacteria), Bacteroidota (Bacteroidetes), Bacillota (Firmicutes), Fusobacteriota (Fusobacteria), Pseudomonadota (Proteobacteria), Saccharibacteria (TM7), and Spirochaetota (Spirochaetes). Within CPR, more than 70 phyla exist, including Saccharibacteria (TM7), ‘Candidatus Absconditabacteria’ (SR1), and ‘Candidatus Gracilibacteria’ (GN02), all commonly detected in the human oral microbiome.9,10
Saccharibacteria are particularly noteworthy due to their high prevalence in the oral cavity and association with mucosal diseases.11 The first cultured CPR bacterium, Nanosynbacter lyticus strain TM7x, revealed an ultrasmall cell size (200-300 nm), a reduced genome with limited biosynthetic capacity, and an epiparasitic lifestyle reliant on the bacterial host Schaalia odontolytica.12 Six major Saccharibacteria clades (G1-G6) exist in the oral cavity, with all cultivated species belonging to clade G1.13 Long-read sequencing has enabled complete genome reconstruction from saliva metagenomes, producing the first complete genomes from Saccharibacteria clade G6, also known as ‘Candidatus Nanogingivalaceae’ and HMT-870.7 Despite numerous associations with disease, Saccharibacteria’s physiology and pathogenesis mechanisms remain poorly understood, though their ecological roles within oral microbiome are illustrated in Figure 1 and some studies report they can reduce bone loss caused by Actinobacteria in mouse models of periodontal disease.5,13 Saccharibacteria, Originally TM7, are widespread and inferred to rely on environmental or host-derived metabolic compounds.23,24
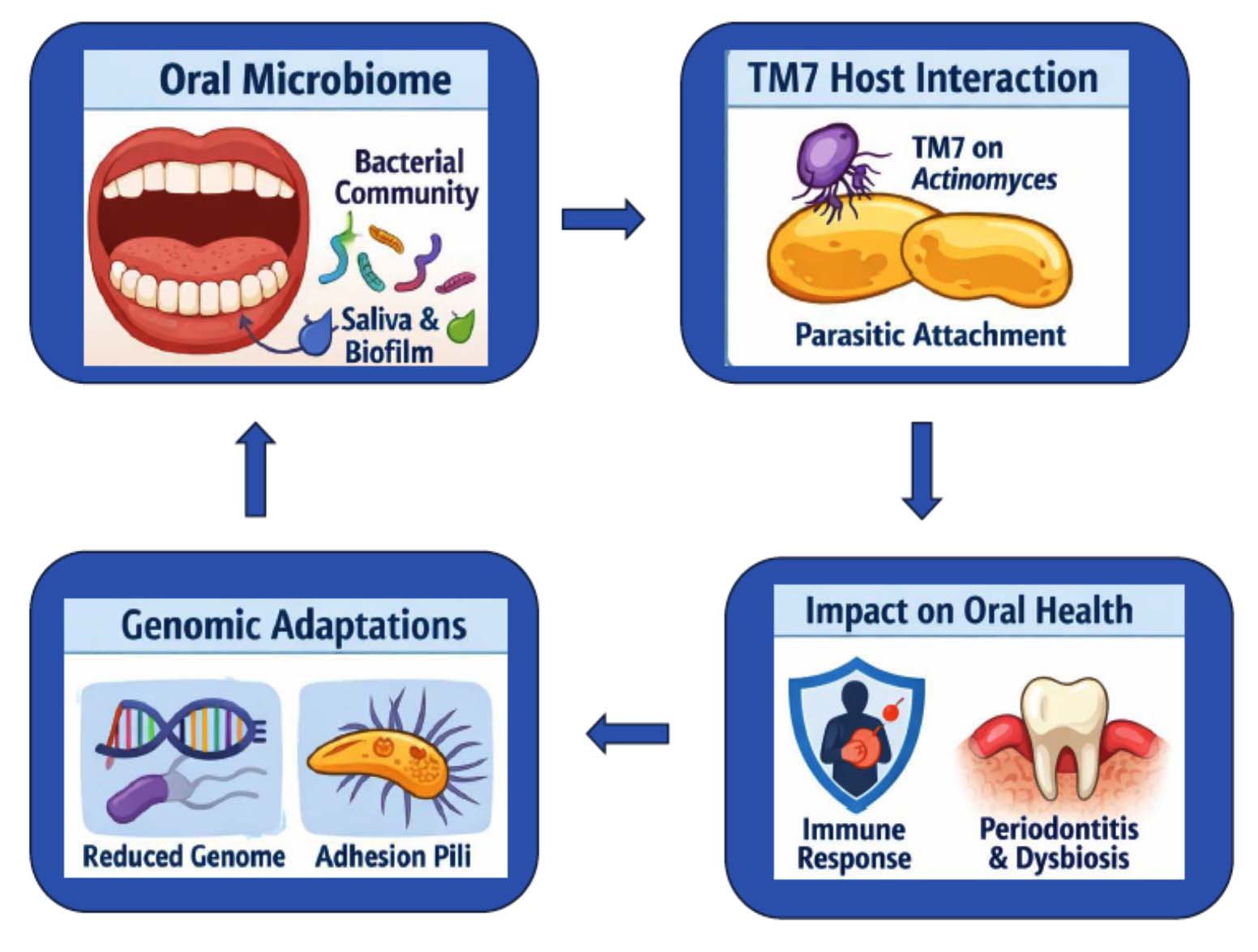
Figure 1. Ecological Roles of TM7 in the Oral Microbiome23
Several species have been cultured as obligate ectoparasites on Actinobacteria.12,24 Using targeted reverse-genomics, “Cand. Nanosynbacter sp. HMT352” strain KC1 (HMT352-KC1) was co-cultured with Schaalia odontolytica strain ORNL 0103.14,24 TM7 represents the first cultured member of CPR, with ~20 years required to uncover its epiparasitic lifestyle.24 Refer Table 1 for six groups (G1-G6) exist in the oral cavity, with cultivated species only from G1 (Cross). TM7 adapt genomically during transition from environmental to mammalian niches. The oral cavity is a major Saccharibacteria habitat, with higher relative abundance than other niches.12 While increased abundance is linked to periodontitis and gingivitis, some Saccharibacteria species reduce inflammatory bone loss by modulating host pathogenicity.5,20
Table (1): Overview of Candidatus Saccharibacteria species/groups, associated diseases, and supporting evidence from the literature
Species/Group |
Disease Association |
Type of Study |
Cultivation |
Ref. |
|---|---|---|---|---|
TM7x (G1) |
Periodontitis, peri-implantitis |
Co-culture, metagenomics |
Cultivated |
[14] |
G1-G5 (Uncultured) |
Periodontitis |
16S rRNA, qPCR |
Uncultivated |
[4] |
G5 |
Oral Lichen Planus (OLP) |
16S rRNA, salivary microbiome |
Uncultivated |
[15] |
G2, G5 |
Dental caries |
Salivary metagenome |
Uncultivated |
[16] |
G3-G5 |
Halitosis |
16S rRNA from tongue coating |
Uncultivated |
[17] |
TM7-like species |
HIV-associated oral dysbiosis |
Oral microbiome profiling |
Uncultivated |
[18] |
TM7-like phylotypes |
COVID-19-associated oral microbiome changes |
Salivary microbiome |
Uncultivated |
[19] |
TM7-G1, G5 |
Peri-implantitis |
Subgingival microbiota study |
Uncultivated |
[20] |
TM7-like (skin-derived) |
Psoriasis, acne (weak association) |
Skin metagenome analysis |
Uncultivated |
[21] |
TM7-like (oral-gut axis) |
Crohn’s disease, IBD (from oral samples) |
Oral microbiome study |
Uncultivated |
[22] |
Oral manifestations of systemic conditions are frequently observed and often associated with changes in the oral microbiome. Cardiovascular disorders, for instance, may be accompanied by periodontal inflammation, gingival bleeding, and enhanced dental plaque formation, indicating a potential bidirectional relationship between oral and cardiac health.25,26 Oral inflammation and gingival bleeding have been correlated with elevated C-reactive protein levels and endothelial dysfunction, suggesting a bidirectional relationship between oral and cardiac health.27 In autoimmune diseases, such as oral lichen planus, reticular or erosive lesions, burning sensations, and alterations in the oral microbiome, including TM7 enrichment, have been observed.15,28
Similarly, systemic lupus erythematosus (SLE) is associated with oral ulcers, hypo-salivation, and higher TM7 abundance in subgingival plaque, TM7 also been implicated in HIV-associated oral dysbiosis COVID-19-related oral microbiome alterations, suggesting potential role in immune-compromised and infectious conditions.18,19,29 In psoriasis, oral mucosal plaques and erythema, along with potential changes in Saccharibacteria populations, have been reported.30 Rheumatoid arthritis presents with gingival enlargement, xerostomia, aphthous ulcers, and microbiome dysbiosis, including correlations with TM7.31 These findings underscore the complex interplay between systemic diseases and the oral microbiome, highlighting the potential role of Saccharibacteria in oral immune modulation and their possible link to systemic inflammation including associations with oral–gut axis and conditions such as Crohn’s disease inflammatory bowel disease.22,32
Advances in culture-independent methods have revolutionized human microbiome studies, revealing significant uncultured diversity.33 Exploration of this microbial “dark matter” led to new bacterial divisions and archaeal divisions (DPANN).34,35 The first TM7 16S rRNA sequence was described in 1996 and later confirmed by phylogenetic studies and genome reconstructions.34,36 CPR bacteria are widely distributed environmentally and in human niches, with Ca. Saccharibacteria and Ca. Absconditabacteria among the most represented phyla.34,37 TM7 represent a unique group of oral microorganisms that depend on specific bacterial hosts, such as Actinomyces, Schaalia, and Arachnia, for survival.14,38
These bacteria have highly reduced genomes and depend on host-derived nutrients, while specialized features like type IV pili and the arginine deiminase pathway support their attachment and persistence in the oral environment.13,39 TM7 display distinct site-specific distributions, being more abundant in saliva, and they play significant roles in shaping microbial networks, modulating immune responses, and influencing oral health outcomes, including maintenance of microbial balance or contribution to periodontitis and caries.5,40 Advanced methodologies, such as coculture experiments and metagenomic analyses, are crucial to fully understand their ecological roles and potential systemic effects.34,38 This review aims to comprehensively examine the diversity, genomic characteristics, ecological functions, and links to disease of TM7 within the oral microbiome, in order to better understand their role in oral microbial communities and potential impact on human health.
Study design
This study was conducted as a systematic review to evaluate the role of Candidatus Saccharibacteria in the human oral microbiome and its associations with oral and systemic health and disease. The review was guided by the PRISMA framework41 to ensure methodological transparency and reliability. The review protocol was registered with PROSPERO (ID: CRD420251026385).
PICO framework
The review applied the PICO framework to clearly define the research scope. Population (P) included humans with a healthy or diseased oral microbiome. Intervention/Exposure (I/E) encompassed the presence, abundance, or host interactions of TM7. Comparison (C) involved different oral sites, health statuses, or detection methods. Outcome (O) focused on prevalence, diversity, genomic features, microbial interactions, and associations with oral or systemic diseases. Using PICO allowed structured selection and extraction of relevant studies to address the research objectives systematically.
Eligibility criteria
This review included studies published from January 2020 to January 2025 in peer-reviewed journals that investigated Candidatus Saccharibacteria within the oral environment. Eligible studies reported on aspects such as prevalence, diversity, taxonomy, genomic characteristics, or microbial interactions, and those examining links between Saccharibacteria and oral or systemic diseases were also considered. Only full-text articles published in English were accepted. Studies were excluded if they were non-peer-reviewed sources, including conference abstracts, preprints, or dissertations, if they were not in English, if they did not present original data (e.g., reviews, commentaries, or editorials), or if they studied only diseased populations without including a healthy comparison group.
Information sources and search strategy
A comprehensive literature search was conducted in January 2025 across multiple databases, including PubMed, Web of Science, Scopus, MEDLINE, and the Cochrane Library, focusing on studies published between 2020 and 2025. The search strategy combined relevant keywords using Boolean operators, such as “Candidatus Saccharibacteria” or “TM7” or “Candidate Phyla Radiation,” together with terms related to the oral environment, including “oral microbiome,” “oral cavity,” or “oral microbiota,” and disease-related keywords, such as “disease,” “periodontitis,” or “systemic.”
Study selection
The initial search found 932 records. After removing duplicates, two reviewers independently looked through the titles and abstracts of all remaining studies. Studies that met the inclusion requirements moved forward for full-text review and detailed examination. When the two reviewers disagreed, a third reviewer helped settle the differences. The entire process was tracked and presented in a PRISMA flow diagram (Figure 2).
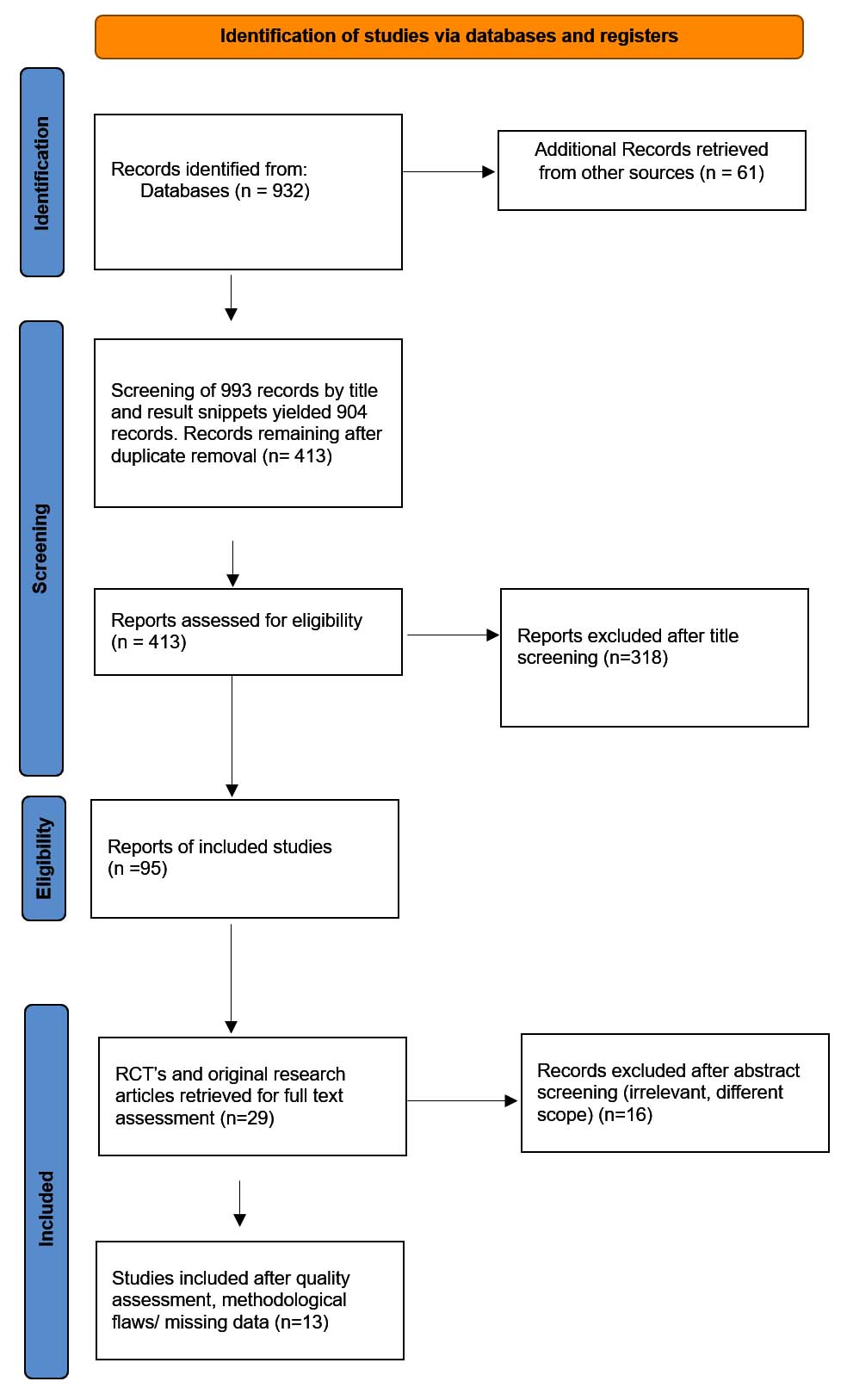
Figure 2. PRISMA Flow Chart of Study Selection
Data extraction and management
Data extraction was carried out independently by two reviewers using a standardized form. The information collected included study details such as author(s), publication year, and location, as well as study design, characteristics of the study population, and the type of sample analysed, including saliva, plaque, or tissue. Details on the analytical methods, such as sequencing techniques and taxonomic tools, and key findings regarding the prevalence, diversity, genomic features, microbial interactions, and associations with oral or systemic diseases were also extracted. All data were subsequently cross-checked to ensure accuracy and consistency.
Risk of bias assessment
The risk of bias for the included studies was evaluated independently by two reviewers using the Cochrane Risk of Bias 2 tool. Each study was assessed across predefined domains and categorized as low risk, some concerns, or high risk. Any differences in judgments were resolved through discussion until agreement was reached. The overall results of this appraisal are summarized in Table 2.
Table (2): Risk of Bias assessment across included studies
Study |
D1 |
D2 |
D3 |
D4 |
D5 |
D6 |
D7 |
Overall |
|---|---|---|---|---|---|---|---|---|
Bachtiar et al.1 |
+ |
− |
+ |
+ |
+ |
+ |
+ |
+ |
Chipashvili et al.5 |
− |
+ |
+ |
+ |
− |
+ |
+ |
− |
Bor et al.14 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Naud et al.34 |
+ |
+ |
− |
+ |
+ |
? |
+ |
− |
He et al.38 |
+ |
+ |
+ |
+ |
+ |
− |
+ |
+ |
Nie et al.39 |
+ |
− |
+ |
+ |
+ |
+ |
+ |
+ |
Wang et al.40 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Papaleo et al.42 |
− |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Murugkar et al.43 |
+ |
+ |
− |
+ |
+ |
+ |
+ |
+ |
Utter et al.44 |
+ |
− |
+ |
+ |
+ |
? |
+ |
− |
Wang et al.45 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
You et al.46 |
− |
+ |
+ |
+ |
+ |
? |
+ |
− |
Naud et al.47 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Note: “+” = low risk of bias, “–” = high risk of bias, and “?” = Insufficient information to make a judgement
The findings of the risk of bias evaluation are summarized in Table 2. Several studies were rated as having a low risk of bias across most domains. However, a considerable number of studies were judged to raise some concerns, particularly regarding allocation concealment and blinding. A few studies were assessed as being at some concerns, mainly due to incomplete outcome data and selective reporting.
Quality assessment
Each included study was evaluated for quality using a standardized appraisal tool designed for microbiome research. Criteria included participant selection, sample size adequacy, sequencing quality, data analysis methods, and reproducibility. Studies were classified as high, moderate, or low quality. Low-quality studies were excluded from the synthesis.
Data synthesis and analysis
Due to considerable variation in study methods, populations examined, and reported outcomes, a meta-analysis could not be performed. Instead, the evidence was synthesized narratively and grouped into four overarching themes: the prevalence and distribution of Saccharibacteria within the oral microbiome, their genomic and functional features that determine host dependence and ecological adaptation, their involvement in oral diseases such as periodontitis and dental caries, and their broader significance in relation to systemic health. Across these themes, recurring patterns were identified, gaps in current understanding were noted, and implications for future lines of investigation were emphasized.
The studies included in this review provide insights into the distribution, diversity, and functional characteristics of TM7 in the oral cavity. They examine its interactions with host bacteria, methods of detection and isolation, and potential roles in oral health and disease. Collectively, the findings illustrate the ecological significance of TM7 and its complex relationships within the oral microbiome. The Table 3 highlights experimental approaches, host interactions, and laboratory-based discoveries on TM7.
Table 3 explains the laboratory studies reveal that TM7 abundance varies across oral sites and correlates with host inflammatory markers, suggesting potential immunomodulatory roles. Coculture experiments with Actinomyces, Schaalia, and Arachnia species demonstrate strong host specificity and sometimes parasitic effects, such as host growth inhibition. Advanced imaging techniques, including FISH, SEM, and confocal microscopy, have visualized TM7 morphology and host attachment behavior. Genomic and metagenomic analyses uncover novel species, metabolic adaptations, and genes involved in host interactions. Cross-site and global screenings indicate that TM7 is predominantly oral-adapted, with occasional systemic colonization. Overall, these laboratory-based findings provide critical insights into TM7 ecology, host relationships, and their potential impact on oral and systemic health.
Table (3): Laboratory Findings on TM7 in Oral and Systemic Microbiomes
Author |
Detection/Isolation Methods |
Advanced Approaches/Techniques |
Host/Coculture Bacteria |
Distinct Laboratory/Experimental Findings |
|---|---|---|---|---|
Bachtiar1 |
Nanopore full-length 16S rRNA sequencing; qPCR for IL-6, CRP, narG, napA |
Correlation of microbial presence with host inflammatory markers; repeated sampling over time; microbial network analysis |
Schaalia odontolytica |
Monitored TM7 abundance across oral niches; verified host-microbe interactions through targeted qPCR. |
Chipashvili5 |
16S rRNA profiling; coculture; FISH; confocal microscopy |
ASV-level microbial profiling using DADA2; high-resolution spectral confocal imaging; murine periodontitis model |
Candidate host bacteria identified by “bait” approach |
Observed TM7 attachment and parasitic dynamics in cocultures; confirmed via FISH and confocal microscopy. |
Bor14 |
Filtration (
| Ultracentrifugation and selective filtration to enrich ultrasmall cells; host range determination; genome-based metabolic profiling |
Actinomyces spp., Pseudopropioni- bacterium propionicum |
Successfully isolated new TM7 strains; detailed morphology via SEM; confirmed host specificity in vitro. |
Naud34 |
PCR, RT-PCR, shotgun metagenomics, SEM |
Combined molecular and ultrastructural approaches; metagenome reconstruction from diverse samples |
Not applicable |
Confirmed ultrasmall TM7 morphology; revealed ecological distribution and enrichment across human body sites. |
He38 |
Metagenomic reconstruction; genome assembly and clustering |
Comparative genomic analysis of over s 2,000 TM7 genomes; identification of metabolic and host-interaction gene |
Not applicable |
Reconstructed 2,041 TM7 genomes; highlighted metabolic potential and adaptive host-associated traits. |
Nie39 |
High-throughput 96-well coculture with 76 basibionts; qPCR; FISH; genome sequencing |
Large-scale coculture screening; genome- based strain characterization; FISH visualization |
Multiple basibionts including Schaalia and Actinomyces spp. |
Isolated 38 TM7 strains; mapped host interactions; characterized phenotypic diversity. |
Wang S40 |
Saliva vs buccal 16S rRNA sequencing (Illumina NovaSeq); QIIME2 |
Oral site-specific microbiome analysis; alpha and beta diversity metrics; co-occurrence network evaluation |
Not applicable |
Mapped TM7 distribution across oral sites; analyzed relative abundance and co-occurrence with other bacteria. |
Papaleo42 |
16S vs 23S rRNA qPCR; metagenomic sequencing |
Developed highly sensitive 23S rRNA qPCR; integrated with metagenomic analysis |
Not applicable |
Demonstrated increased sensitivity in TM7 detection; uncovered previously undetected lineages. |
Murugkar43 |
Coculture with Arachnia & Actinomyces; 16S rRNA; genome sequencing |
Assessment of host specificity; evaluation of coculture stability; long-term culture maintenance |
Arachnia propionica, Actinomyces spp. |
Evaluated stability of TM7-host cocultures; studied effects on host growth and morphology. |
Utter44 |
Coculture; PCR; FISH; growth assays; comparative genomics |
Gradient infection experiments; correlation with host genotypes; pangenomic analysis |
37 Actinomyces strains |
Documented TM7x-triggered growth crashes; linked host susceptibility to genotype; visualized parasitic behavior. |
Wang45 |
EMP global saliva dataset; standardized 16S workflow |
Global meta-analysis of TM7-related taxa; microbial network ecology analysis |
Not applicable |
Identified core TM7-associated taxa worldwide; revealed ecological interactions in oral microbiomes. |
You46 |
16S rRNA sequencing (Illumina); QIIME2; DADA2 |
Age-stratified microbial analysis; taxonomic co-occurrence evaluation |
Not applicable |
Correlated TM7 abundance with caries-associated taxa; highlighted age-dependent microbial patterns. |
Naud47 |
PCR; Nanopore; Illumina; coculture; SEM |
Genome sequencing of fecal TM7 isolates; SEM imaging; host specificity verification |
Schaalia odontolytica, Arachnia propionica |
Isolated Ca. M. massiliensis and Ca. M. timonensis; characterized morphology and metabolism; confirmed host specificity. |
Table (4): Summary of Key findings on Saccharibacteria from Recent laboratories and Clinical studies
Authors |
Study Design/Sample Sources |
Sample/Population |
Key Findings on Saccharibacteria |
Host Interaction Observations |
Clinical/Health Implications |
Ref. |
|---|---|---|---|---|---|---|
Bachtiar et al.1 |
Longitudinal clinical study (mini-implants) |
8 healthy adults; saliva, gingival crevicular fluid (GCF), peri-implant crevicular fluid at baseline, 1 week, 4 weeks |
Saccharibacteria levels decreased over time; sensitive to ecological stress around implants |
Inversely correlated with IL-6; antagonistic toward P. gingivalis, T. denticola, F. nucleatum |
May play a protective or immunomodulatory role during inflammatory events |
[1] |
Chipashvili et al.5 |
Clinical plaque sampling + mouse model |
Patients with periodontitis and mice |
Confirmed TM7 in periodontal plaque; host-specific growth limitations |
Cocultures increased inflammatory cytokines and bone loss in mice |
TM7 may worsen periodontitis progression |
[5] |
Bor et al.14 |
Coculture isolation |
44 adults; saliva and plaque |
Three new strains isolated (BB001, AC001, PM004) |
Strong host specificity; parasitic growth can reduce host viability |
Demonstrates methods to maintain stable TM7 cultures in vitro |
[14] |
Naud et al.34 |
Cross-body site screening |
Oral cavity, gut, breast milk, blood, heart valves |
Oral cavity harbors highest TM7 abundance (95% detection by RT-PCR); ultrasmall cocci
| Most enrichment oral/ respiratory; systemic presence uncommon |
TM7 mainly oral-adapted; occasional systemic colonization |
[34] |
He et al.38 |
Metagenomic reconstruction |
>4,000 oral samples (China + public datasets) |
293 novel species; six new genera; five new families; genomes highly reduced metabolically |
Presence of ADS and type IV pili genes → resistance to acidic stress and enhanced adhesion |
TM7 diversity linked to oral diseases and systemic conditions such as rheumatoid arthritis |
[38] |
Nie et al.39 |
High-throughput isolation |
16 patients with periodontitis; saliva |
38 TM7 strains isolated; broad host compatibility |
Phenotypic variability; some induce host growth crash |
High strain-level diversity → variable effects on pathogenicity |
[39] |
Wang S et al.40 |
Oral site comparison |
50 healthy adults |
Saccharibacteria 2.86% in saliva vs 0.29% in buccal (P
| Saliva microbiome more diverse and deterministic |
Oral site strongly influences TM7 distribution |
[40] |
Papaleo et al.42 |
Comparison of diagnostic methods |
Oral samples |
23S qPCR more sensitive; detected more Saccharibacteria |
Novel lineages identified in the oral microbiome |
Standardized detection methods important; linked to allergy susceptibility in children |
[42] |
Murugkar et al.43 |
Coculture and host specificity |
Oral samples from multiple sites |
HMT-488 and HMT-955
successfully cocultured |
Strong host specificity; some cocultures stable, others unstable |
Highlights TM7’s ecological role in shaping bacterial communities |
[43] |
Utter et al.44 |
Host range investigation |
37 Actinomyces strains tested |
TM7x limited to clade-2 Actinomyces |
Growth crash phenotypes associated with host susceptibility |
Host genetics influence TM7 resistance (e.g., glycosyltransferases, ADI) |
[44] |
Wang et al.45 |
EMP global saliva dataset |
404 saliva samples |
14 core OTUs including Saccharibacteria; regional variation observed |
Co-occurrence networks highlight interactions |
TM7 forms part of oral microbiome core taxa; less diversity than gut or skin |
[45] |
You et al.46 |
Pediatric caries study |
102 children (ages 3-8); plaque and saliva |
Saccharibacteria consistently correlated with caries severity; enriched in diseased sites |
Positively associated with Fusobacterium; negatively with Streptococcus |
May contribute to caries development; microbial beta diversity varies with age |
[46] |
Naud et al.47 |
Fecal TM7 study |
2 HIV-positive patients |
Identified “Candidatus Massiliensis massiliensis” and “Candidatus M. timonensis” |
Successfully cocultured with Actinomyces and Schaalia |
Suggests TM7 host range can extend beyond oral cavity into gut |
[47] |
Table 4, describes recent investigations have provided valuable insights into TM7 biology and its interactions with host bacteria. Longitudinal clinical studies reveal site-specific abundance changes and potential immunomodulatory roles. Coculture experiments demonstrate strong host specificity, parasitic growth patterns, and effects on host viability, influencing microbial community dynamics. Metagenomic analyses have identified novel species and genera, highlighted metabolic adaptations, and pinpointed genes involved in adhesion and stress resistance. Cross-body site screenings confirm that TM7 is predominantly oral-adapted, with rare systemic presence. Collectively, these studies emphasize TM7’s ecological significance, host-dependent behavior, and implications for oral and systemic health.
Candidatus Saccharibacteria represents a key yet understudied member of the human oral microbiome. Evidence from multiple investigations indicates that TM7 is primarily adapted to the oral environment, exhibits strict host dependence, and influences microbial community composition and host interactions in nuanced ways. Collectively, these studies provide insights into the ecological, genomic, and functional characteristics of TM7 relevant to oral health and disease.
Prevalence and Site-Specific Colonization: TM7 is widely detected in the oral cavity, although its abundance varies across different sites. Study observed a temporal decline in TM7 populations in saliva and peri-implant crevicular fluid during orthodontic treatment, implying that environmental or clinical interventions can modulate its prevalence.1 Study reported higher TM7 presence in saliva compared with the buccal mucosa, highlighting site-specific colonization patterns.40 Broad surveillance studies support these findings: Naud et al. reported detection of TM7 in approximately 95% of oral samples, whereas its presence in faecal matter, breast milk, urine, and blood remained limited.34 Methodological approaches also influence observed prevalence; demonstrated that conventional 16S rRNA-based assays tend to underestimate TM7, whereas 23S rRNA sequencing and metagenomic approaches provide more accurate detection.42
Host Dependency and Coculture Dynamics: TM7 relies on specific oral bacteria for growth, emphasizing its obligate host dependence. Study successfully cocultured novel TM7 strains (BB001, AC001, PM004) with Actinomyces and Pseudopropionibacterium hosts, while Murugkar et al.43 confirmed that only select Actinomyces spp. and Arachnia propionica can serve as basibionts. Study highlighted intra-species variation, showing that some TM7 strains trigger host growth-crash while others persist without damaging the host.39 Also further reported strain-dependent host responses, including transient growth inhibition in permissive hosts and stable colonization in resistant hosts.44 These findings collectively underscore the ecological influence of TM7 as mediated by host-specific interactions, which can modulate overall oral microbial composition.
Advanced Techniques for TM7 Detection and Analysis: Several studies employed advanced methodologies to investigate TM7, but their approaches varied significantly. In Bachtiar et al.,1 study integrated host inflammatory markers with Nanopore sequencing and network analysis, whereas Papaleo et al. focused on improving detection sensitivity using 23S rRNA qPCR combined with metagenomic sequencing, highlighting methodological differences in microbial detection.42 In contrast, Bor et al.14 and Nie et al.39 emphasized coculture-based strategies paired with FISH and SEM imaging to visualize TM7-host interactions and phenotypic diversity, while Utter et al.44 introduced gradient infection assays and pangenomic analyses to link host genotype with susceptibility. Studies such as Chipashvili et al.5 and Naud et al.34 combined high-resolution imaging with computational or genomic approaches, providing a more integrated view of TM7 morphology and host specificity.
Microbial interactions and community integration
TM7 participates in complex interspecies networks within the oral microbiome. Study described antagonistic interactions between TM7 and key periodontopathogens, while nitrate-reducing bacterial populations remained stable, suggesting a potential buffering role.1 TM7 also serves as a connector in microbial networks, associating with Fusobacterium nucleatum and other members of the red complex to influence community structure. He et al. emphasized that, despite reduced metabolic potential, TM7 retains functional mechanisms such as type IV pili and arginine deiminase pathways that facilitate host adherence and survival.38 Study reported that saliva microbiomes exhibit deterministic assembly with structured TM7-mediated interspecies relationships, whereas buccal mucosa microbiomes display more stochastic patterns.45 He demonstrated that TM7-host complexes can exacerbate inflammatory responses and bone resorption in vivo, illustrating context-dependent ecological roles.5
Genomic Features and Functional Adaptations: Genomic analyses reveal that TM7 harbors highly reduced genomes, typically under 800 kbp, with limited biosynthetic capabilities, confirming its obligate dependence on hosts.14,38,39 Despite this reduction, TM7 exhibits specialized adaptations for survival in oral niches, including type IV pili for adhesion and arginine deiminase systems for acid tolerance. The author identified substantial genetic heterogeneity across TM7 strains, including accessory genes that may influence host specificity, infection dynamics, and phenotypic outcomes.39 Also further highlighted that host glycosylation genes and arginine deaminase pathway components contribute to susceptibility to TM7 colonization, illustrating the genomic interplay between parasite and host.44
Phenotypic variability and ecological implications
TM7 exhibits strain-dependent phenotypic variation with ecological relevance. Bor et al. observed distinct morphologies and host-induced modifications, such as elongation and swelling.14 Study reported differences in growth-crash induction and infection timing among strains,39 while Utter et al. described unique attachment patterns to specific hosts.44 These phenotypic differences suggest that TM7 can exert selective pressures on host populations, modulating microbial community structure and influencing the balance between symbiosis and pathogenicity.
Host immune modulation
TM7 interactions with the host immune system appear context-dependent. Study noted inverse correlations between TM7 abundance and IL-6 levels following mini-implant placement, indicating potential immunomodulatory roles. Conversely, observed that TM7-host complexes increased pro-inflammatory cytokine expression in gingival tissues, contributing to alveolar bone loss. These findings imply that TM7 may exert protective or deleterious effects depending on host, strain, and ecological context.
Methodological considerations
Detection and characterization of TM7 are influenced by methodological approaches. High-resolution techniques, including ultrafiltration, anaerobic coculture, long-read metagenomics, and fluorescence in situ hybridization, allow precise identification and functional analysis.1,14,39 In contrast, reliance on conventional 16S rRNA-based methods may underestimate TM7 prevalence and obscure strain-level diversity.43 Integrative multi-modal approaches are essential for capturing TM7 dynamics accurately in the human oral ecosystem.
Candidatus Saccharibacteria (TM7) emerges as a highly specialized and ecologically significant member of the oral microbiome. Its strict dependence on selects host bacteria and site-specific abundance highlight a finely tuned ecological role, while strain-level phenotypic and genomic diversity points to its capacity to modulate microbial community dynamics and influence host-microbe interactions. TM7’s context-dependent effects on the immune system suggest it may contribute to both protective and pathogenic processes in oral tissues.
The current understanding of TM7 remains limited by methodological constraints, with detection and functional characterization strongly influenced by the approaches used, such as coculture, metagenomics, and FISH. Moreover, the diversity of TM7-host interactions, metabolic adaptations, and ecological strategies remains largely unexplored. A deeper investigation into these aspects could reveal critical insights into biofilm formation, microbial competition, and the balance between health and disease. Expanding knowledge on TM7’s functional roles may also provide novel perspectives on microbial ecology and its contribution to oral and systemic pathologies, making focused research both timely and necessary.
Limitations
This review is limited by the small number of eligible studies (13 in total) and the considerable heterogeneity in study designs, ranging from coculture experiments to metagenomic analyses. Differences in detection methods (16S vs. 23S rRNA sequencing, qPCR, metagenomics) may have influenced prevalence estimates. Most studies were either cross-sectional or laboratory-based, with limited longitudinal or clinical cohort data, reducing the ability to infer causal relationships. In addition, geographic representation was narrow, restricting generalizability across diverse populations.
ACKNOWLEDGMENTS
The authors would like to acknowledge Sri Balaji Vidyapeeth Deemed to be University, Puducherry, for their support.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
VA conceptualized the study and performed project administration. VN collected resources and supervised the study. DR performed data curation and formal analysis. SM performed software, validation, wrote and revised the manuscript. SDA performed visualization. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
ETHICS STATEMENT
Not applicable.
- Bachtiar BM, Tahapary DL, Fath T, et al. Saccharibacteria (TM7) in saliva and subgingival microbiome as a predictor for gingivitis in individuals with type 2 diabetes evaluated by qPCR. Front Dent Med. 2025;6:1550936.
Crossref - Chen T, Yu WH, Izard J, Baranova OV, Lakshmanan A, Dewhirst FE. The human oral microbiome database: a web accessible resource for investigating oral microbe taxonomic and genomic information. Database (Oxford). 2010;2010:baq013.
Crossref - Camanocha A, Dewhirst FE. Host-associated bacterial taxa from Chlorobi, Chloroflexi, GN02, Synergistetes, SR1, TM7, and WPS-2 phyla/candidate divisions. J Oral Microbiol. 2014;6: 25468.
Crossref - Brinig MM, Lepp PW, Ouverney CC, Armitage GC, Relman DA. Prevalence of bacteria of division TM7 in human subgingival plaque their association with disease. Appl Environ Microbiol. 2003;69(3):1687-1694.
Crossref - Chipashvili O, Utter DR, Bedree JK, et al. Episymbiotic Saccharibacteria suppresses gingival inflammation and bone loss in mice through host bacterial modulation. Cell Host Microbe. 2021;29(11):1649-1662.e7.
Crossref - Mukherjee C, Moyer CO Steinkamp HM, et al. Acquisition of oral microbiota is driven by environment, not host genetics. Microbiome. 2021;9(1):54.
Crossref - Baker JL, Mark Welch JL, Kauffman KM, McLean JS, He X. The oral microbiome: diversity, biogeography and human health. Nat Rev Microbiol. 2024;22(2):89-104.
Crossref - Baker JL, Bor B, Agnello M, Shi W, He X. Ecology of the oral microbiome: beyond bacteria. Trends Microbiol. 2017;25(5):362-374.
Crossref - Shaiber A, Willis AD, Delmnt TO, et al. Functional and genetic markers of niche partitioning among enigmatic members of the human oral microbiome. Genome Biol. 2020;21(1):292.
Crossref - Jaffe AL, Thomas AD, He C, et al. Patterns of gene content and co-occurrence constrain the evolutionary path toward animal association in candidate phyla radiation bacteria. mBio. 2021;12(4): e0052121.
Crossref - McLean JS, Bor B, Kems KA, et al. Acquisition and adaptation of ultra-small parasitic reduced genome bacteria to mammalian hosts. Cell Rep. 2020;32(3):107939.
Crossref - He X, McLean JS, Edlund A, et al. Cultivation of a human-associated TM7 phylotype reveals a reduced genome and epibiotic parasitic lifestyle. Proc Natl Acad Sci USA. 2015;112(1):244-249.
Crossref - Tian J, Utter DR, Cen L, et al. Acquisition of the arginine deiminase system benefits epiparasitic Saccharibacteria and their host bacteria in a mammalian niche environment. Proc Natl Acad Sci USA. 2022;119(5):e2114909119.
Crossref - Bor B, Collins AJ, Murugkar PP, et al. Insights obtained by culturing Saccharibacteria with their bacterial hosts. J Dent Res. 2020;99(6):685-694.
Crossref - Jungpraditphol I, Sutthiboonyapan P, Khamwachirapitak C, et al. Shotgun Metagenomics of Biofilm Microbiome in Oral Lichen Planus with Desquamative Gingivitis. Oral Dis. 2025;31(11):3069-3080.
Crossref - Belda-Ferre P, Alcaraz LD, Cabrera-Rubio R, et al. The oral metagenome in health and disease. ISME J. 2012;6(1):46-56.
Crossref - Takeshita T, Suzuki N, Nakano Y, et al. Discrimination of the oral microbiota associated with high hydrogen sulfide and methyl mercaptan production. J Clin Microbiol. 2012;50(2):319-326.
Crossref - Kistler JO, Arirachakaran P, Poovorawan Y, Dahlén G, Wade WG. The oral microbiome in human immunodeficiency virus (HIV)-positive individuals.
J Med Microbiol. 2015;64(9):1094-1101.
Crossref - Rafiqul Islam SM, Foysal MJ, Hoque MN, et al. Dysbiosis of Oral and Gut Microbiomes in SARS-CoV-2 Infected Patients in Bangladesh: Elucidating the Role of Opportunistic Gut Microbes. Front Med (Lausanne). 2022; 9:821777.
Crossref - Al-Ahmad A, Muzafferiy F, Anderson AC, et al. Shift of microbial composition of peri-implantitis-associated oral biofilm as revealed by 16S rRNA gene cloning. J Med Microbiol. 2018;67(3):332-340.
Crossref - Oh J, Byrd AL, Park M, Kong HH, Segre JA. Temporal stability of the human skin microbiome. Cell. 2016;165(4):854-866.
Crossref - Schmidt TS, Hayward MR, Coelho LP, et al. Extensive transmission of microbes along the oral–gut axis. Cell Host Microbe. 2019;26(3):421-432.e5.
Crossref - Bor B, Bedree JK, Shi W, McLean JS, He X. Saccharibacteria (TM7) in the human oral microbiome. J Dent Res. 2019;98(5): 500-509.
Crossref - Cross KL, Klingeman DM, Podar M. Complete genome sequence of human oral Saccharibacterium “Candidatus Nanosynbacter sp. HMT352” strain KC1. Microbiol Resour Announc. 2022;11(2):e01205-21.
Crossref - Tonetti MS, Van Dyke TE, Working Group 1 of the Joint EFP/AAP Workshop. Periodontitis and atherosclerotic cardiovascular disease: consensus report of the Joint EFP/AAP Workshop on Periodontitis and Systemic Diseases. J Periodontol. 2013;84(4 Suppl):S24-S29.
Crossref - Lockhart PB, Bolger AF, Papapanou PN, et al. Periodontal disease and atherosclerotic vascular disease: does the evidence support an independent association? Circulation. 2012;125(20):2520-2544.
Crossref - Humphrey LL, Fu R, Buckley DI, Freeman M, Helfand M. Periodontal disease and coronary heart disease incidence: a systematic review and meta-analysis. J Gen Intern Med. 2008;23(12):2079-2086.
Crossref - Al-Hashimi I, Schifter M, Lockhart PB, et al. Oral lichen planus and oral lichenoid lesions: diagnostic and therapeutic considerations. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2007;103(1 Suppl): S25.e1-S25.e12.
Crossref - Katz-Agranov N, Zandman-Goddard G. The microbiome and systemic lupus erythematosus. Immunol Res. 2017;65(2):432-437.
Crossref - de Pablo P, Chapple IL, Buckley CD, Dietrich T. Periodontitis in systemic rheumatic diseases. Nat Rev Rheumatol. 2009;5(4):218-224.
Crossref - Gao L, Cheng Z, Zhu F, Bi C, Shi Q, Chen X. The oral microbiome and its role in systemic autoimmune diseases: a systematic review of big data analysis. Front Big Data. 2022;5:927520.
Crossref - Sanz M, Del Castillo AM, Jepsen S, et al. Periodontitis and cardiovascular diseases: consensus report. J Clin Periodontol. 2020;47(3):268-288.
Crossref - Lok C. Mining the microbial dark matter. Nature. 2015;522(7556):270-273.
Crossref - Naud S, Valles C, Abdillah A, et al. Preliminary landscape of Candidatus Saccharibacteria in the human microbiome. Front Cell Infect Microbiol. 2023;13:1195679.
Crossref - Rinke C, Schwientek P, Sczyrba A, et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature. 2013;499(7459):431-437.
Crossref - Hugenholtz P, Goebel BM, Pace NR. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J Bacteriol. 1998;180(18):4765-4774.
Crossref - Ji Y, Zhang P, Zhou S, Gao P, Wang B, Jiang J. Widespread but poorly understood bacteria: candidate phyla radiation. Microorganisms. 2022;10(11):2232.
Crossref - He W, Liang H, Li W, et al. Revealing an unprecedented diversity of episymbiotic Saccharibacteria in a high-quality genome collection. NPJ Biofilms Microbiomes. 2024;10(1):153.
Crossref - Nie J, Utter DR, Kerns KA, et al. Strain-level variation and diverse host bacterial responses in episymbiotic Saccharibacteria. mSystems. 2022;7(2):e0148821.
Crossref - Wang S, Song F, Gu H, et al. Comparative evaluation of the salivary and buccal mucosal microbiota by 16S rRNA sequencing for forensic investigations. Front Microbiol. 2022;13:777882.
Crossref - Page MJ, McKenzie JE, Bossuyt PM, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. 2021;372:n71. Published 2021 Mar 29.
Crossref - Papaleo S, Nodari R, Sterzi L, et al. Comparison of qPCR protocols for quantification of “Candidatus Saccharibacteria,” belonging to the candidate phyla radiation, suggests that 23S rRNA is a better target than 16S rRNA. PLoS One. 2024;19(12):e0310675.
Crossref - Murugkar PP, Collins AJ, Chen T, Dewhirst FE. Isolation and cultivation of candidate phyla radiation Saccharibacteria (TM7) bacteria in coculture with bacterial hosts. J Oral Microbiol. 2020;12(1):1814666.
Crossref - Utter DR, He X, Cavanaugh CM, McLean JS, Bor B. The Saccharibacterium TM7x elicits differential responses across its host range. ISME J. 2020;14(12):3054-3067.
Crossref - Wang J, Feng J, Zhu Y, Li D, Wang J, Chi W. Diversity and biogeography of human oral saliva microbial communities revealed by the Earth Microbiome Project. Front Microbiol. 2022;13:931065.
Crossref - You Y, Yin M, Zheng X, et al. Saccharibacteria (TM7), but not other bacterial taxa, are associated with childhood caries regardless of age in a South China population. Peer J. 2023;11: e15605.
Crossref - Naud S, Caputo A, Anani, et al. Genomic description and prevalence of two new Candidatus Saccharibacteria species from the human gut in different samples and countries. New Microbes and New Infections. 2023;54: 101153.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.