


The Oropouche virus (OROV) is an emerging arbovirus grouped under Orthobunyavirus genus of Peribunyaviridae family, first identified in Trinidad and Tobago in 1955, and re-emerged as a serious public health concern in the Central and South Americas. Recently, numbers of OROV cases are on the rise with the 2024 outbreak documenting over 12,000 cases across endemic and newly affected regions, including first-time detections in Caribbean nations and imported cases in the United States, Canada, and Italy. The OROV has caused over 5,00,000 cases globally, with substantial underreporting owing to clinical similarities with Zika, chikungunya and dengue. The virus is mainly transmitted by midges viz. Culicoides paraensis (Goeldi), though alternative vectors such as Culex quinquefasciatus Say and Aedes serratus (Theobald) have also been identified. Clinical manifestations range from acute febrile illness with biphasic fever in approximately 60% of cases to severe complications including haemorrhagic symptoms along with neurological sequelae such as meningitis, encephalitis, and Guillain-Barre syndrome. The virus exhibits complex transmission cycles (urban and sylvatic) and reassortment-driven evolution. The evidence of vertical transmission has emerged, with documented cases of fetal microcephaly, stillbirths, and congenital malformations. Pathogenesis involves modulation of host innate immunity, particularly interferon responses. The virus demonstrates neuroinvasive potential through blood-brain barrier penetration and has been shown to persist in various tissues. Diagnostic challenges arise from non-specific clinical signs and limited access to specialized laboratory tests. At present, no specific treatment or vaccines against the OROV are available and management limited to supportive care. However, promising vaccine candidates such as live-attenuated, inactivated, and recombinant protein-based platforms are undergoing preclinical and clinical evaluation, offering hope for future preventive strategies against this neglected tropical disease.
Oropouche Virus, Epidemiology, Vertical Transmission, Neurological Complications, Diagnosis, Vaccines
The Oropouche virus (OROV) is an important emerging arbovirus in the genus Orthobunyavirus under the order Bunyavirales and family Peribunyaviridae. The OROV is characterised by negative-sense, single-stranded RNA genome encased in a spherical lipid sheath. The virus was recognized in the year 1955 during an outbreak of febrile disease in Trinidad and Tobago. Since then, the OROV has caused numerous outbreaks in Central and South America, particularly in Peru, Brazil and Panama.1,2 To date more than 5,00,000 cases were reported globally, though the actual number of cases might be higher owing to under diagnosis and misdiagnosis, as OROV symptoms often mimic other diseases caused by arboviruses such as Zika, dengue, and chikungunya.3,4
Although Culicoides paraensis (Goeldi) is recognized as the dominant vector for OROV in forested and aquatic habitats, the epidemiological landscape also involves Culicidae species.5 Notably, both Culex quinquefasciatus Say and Aedes serratus (Theobald) have been identified as secondary vectors for the virus.6 Infection with OROV typically results in acute febrile illness characterized by symptoms such as arthralgia, fever, headache and myalgia. In approximately 60% of cases recurrent febrile illness is observed in which symptoms may reappear during one to two weeks after recovery, a phenomenon known as biphasic fever. Severe cases may involve hemorrhagic symptoms, such as epistaxis and gingival bleeding, or neurological complications, including meningitis and encephalitis and peri-infectious Guillain-Barre syndrome.7,8 The OROV has the ability to invade the nervous system in animal models, providing critical insights into its potential to cause neurological complications in humans.9 These conditions highlight the capacity of the virus to cause significant neurological damage, particularly in severe or complicated infections.
Serological evaluation of OROV-infected patients has demonstrated the presence of specific IgM and IgG antibodies via neutralization assays. Furthermore, molecular detection of OROV RNA using quantitative reverse transcription PCR (RT-qPCR) has revealed viral persistence in various biological matrices including urine, plasma and whole blood for up to 31 days following the initial presentation of symptoms.10 Despite its significant public health impact, no specific treatment and licensed vaccines are available to date for OROV. Management of the patients mainly focuses on symptomatic relief and supportive care. However, recent advances in vaccine development have shown promising protection with live attenuated virus, inactivated virus and recombinant protein-based vaccines, which are undergoing preclinical and clinical trials.11
Epidemiology
Since its discovery in 1955, multiple epidemic waves of OROV transmission have occurred across rural and urban landscapes in Central and South America. A landmark epidemiological event occurred in 1961 in Belém, Brazil, where an estimated 11,000 individuals were infected. Taxonomically, OROV belongs to the Simbu serogroup, a cluster of 25 viruses organized into two primary phylogenetic lineages: subclade A, containing the Manzanilla and Oropouche viruses, and subclade B, which encompasses the Simbu, Akabane, Sathuperi, Shamonda, and Shuni viruses.12,13
In 2024, the Oropouche virus epidemic landscape has significantly expanded worldwide with the World Health Organization (WHO) documenting more than 10,000 cases across both endemic and newly affected regions. Notably, the virus has been identified for the first time in several Caribbean nations, including Barbados, Guyana, and Cuba, with Cuba reporting its earliest outbreak in May 2024.14 Moreover, imported cases have been detected in United States, demonstrating that this geographic expansion has transcended the Americas, i.e. 108 confirmed cases linked to travel from Cuba, Canada, and Italy have been reported (Table).15-18 While WHO currently evaluates the public health risk as high for affected regions in the Americas, the global risk assessment remains classified as low. The surveillance data underscores the dynamic nature of OROV transmission and highlights the significance of enhanced monitoring, particularly due to the capacity of OROV for geographic expansion through travellers. The emergence in non-endemic areas warrants heightened vigilance in clinical and public health sectors to ensure timely detection and OROV related response to potential outbreaks.
Table: Epidemiological data regarding Oropouche virus incidence across Pan America since 2024
No. |
Country name |
No. of OROV cases (2025*) |
No. of OROV cases (2024) |
Death/Complications (2025) |
|---|---|---|---|---|
1 |
Brazil |
11888 |
13785 |
5 |
2 |
Panama |
501 |
16 |
1 |
3 |
Peru |
330 |
1263 |
0 |
4 |
Cuba |
28 |
626 |
4 |
5 |
Colombia |
26 |
74 |
0 |
6 |
Canada |
1 (Imported) |
2 |
0 |
7 |
Chile |
2 (Imported) |
– |
0 |
8 |
Uruguay |
3 (Imported) |
– |
0 |
9 |
USA |
1 (Imported) |
108 |
0 |
10 |
Guyana |
1 |
3 |
0 |
11 |
Venezuela |
5 |
– |
0 |
12 |
Barbados |
– |
2 |
0 |
13 |
Bolivia |
– |
356 |
0 |
14 |
Ecuador |
– |
3 |
0 |
15 |
Cayman Island |
– |
1 |
0 |
Total |
12786 |
16239 |
10 |
*Indicates data up to 13 Aug 2025 (PAHO, 2025)
A recent outbreak in non-endemic regions, such as French Guiana and Colombia, further highlights the potential of OROV to spread beyond its traditional geographic range. In 2020, an outbreak of dengue-like illness in French Guiana was attributed to OROV, with 82% of tested patients showing evidence of infection.19 Similarly, phylogenetic analysis of OROV strains from Colombia revealed at least two independent introductions of the virus, underscoring the importance for continuous surveillance and mitigation strategies.20 The timeline of the major occurrence and detection of the OROV was presented in Figure 1.
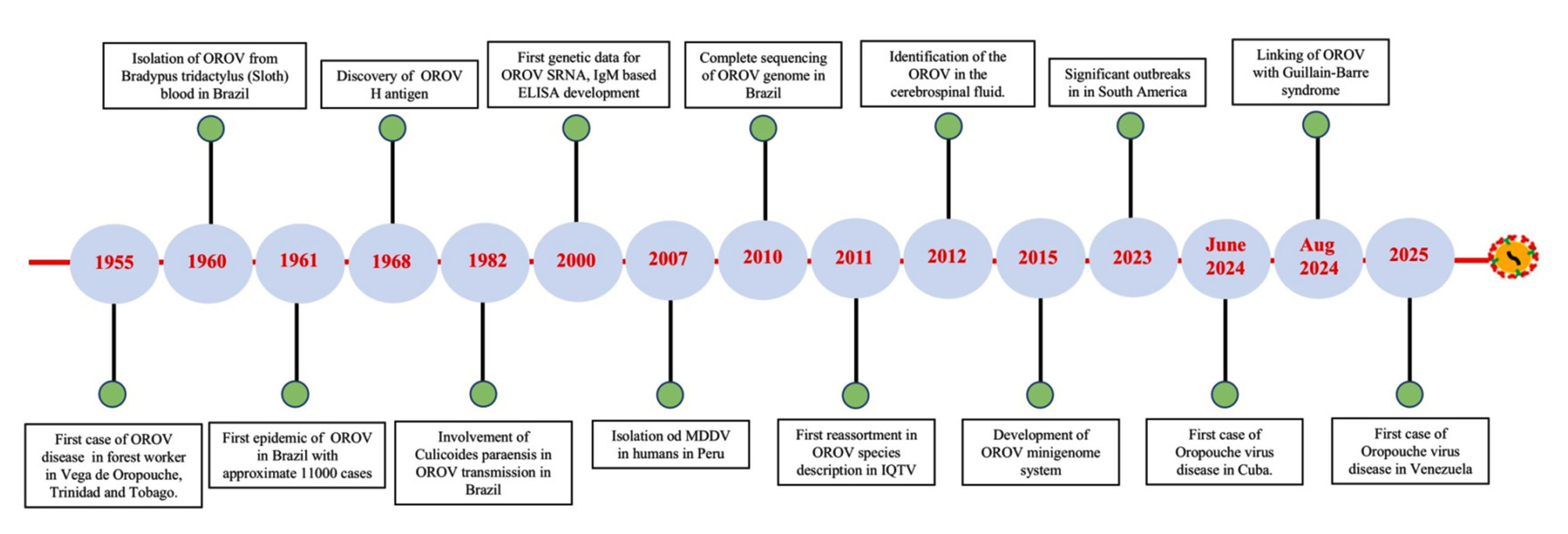
Figure 1. Timeline of the major occurrences and detection of the Oropouche virus
Viral genome
As a member of the Peribunyaviridae family, OROV is classified under genus Orthobunyavirus, which is part of the most extensive group of RNA viruses, comprising over 170 different agents classified into 48 species and 18 serogroups. The virion is morphologically described as spherical in shape and measures 80-120 nm in diameter. The virion is composed of segmented RNA, which is associated with a ribonucleoprotein (RNP) core and is enveloped in a membrane derived from the host cell membrane, which is isolated during the budding process (Figure 2).
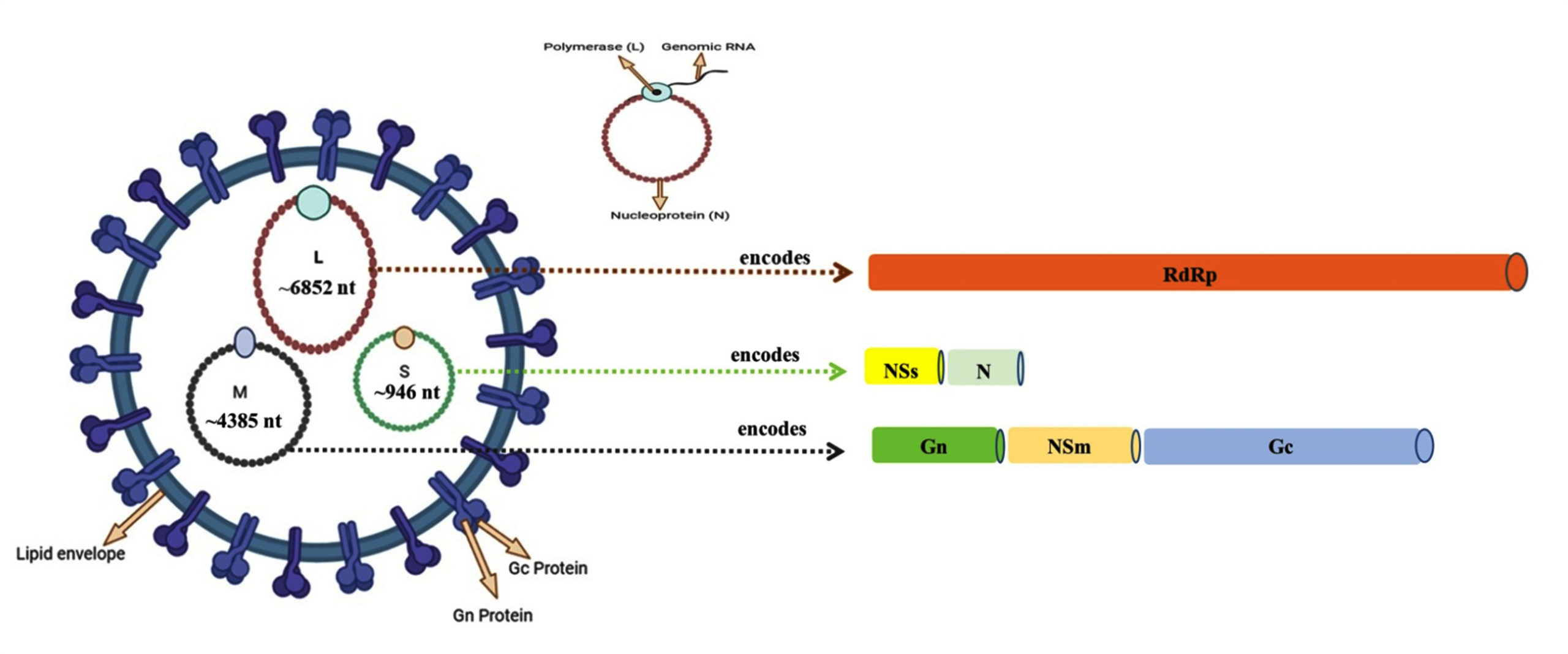
Figure 2. Schematic diagram of Oropouche virus showing a negative-sense, single-stranded RNA genome divided into three segments, i.e. L (Large), M (Medium) and S (Small) encoding six structural and non-structural proteins. The L segment (~6,852 nucleotides) encodes for RNA-dependent RNA polymerase (RdRp), M segment (~4,385 nucleotides) produces two structural glycoproteins (Gn and Gc) and the non-structural NSm protein located between them and the S segment (~946 nucleotides) encodes both the nucleocapsid (N) protein and non-structural NSs protein
Although high-resolution structural data for OROV is not yet available, studies of related Orthobunyavirus suggest OROV virions are approximately 90 nm in diameter with surface projections formed by glycoprotein N (Gn) and glycoprotein C (Gc). The Orthobunyavirus genome possess a negative-sense, single-stranded RNA divided into three segments, i.e. Large (L), Medium (M), and Small (S) encoding six structural and non-structural (NS) proteins. The L segment (~6,852 nucleotides) encodes for the RNA-dependent-RNA polymerase (RdRp) required for synthesis of viral RNA and transcription.21 The M segment (~4,385 nucleotides) produces a polyprotein precursor cleaved into two structural glycoproteins viz. Gn and Gc and non-structural protein encoded by the M (medium) RNA segment (NSm) located between them.4,21 The Gn and Gc are type I transmembrane proteins featuring N-terminal ectodomains that undergo coordinated processing through the host secretory pathway. The Gn contains a small ectodomain (~200 amino acids) with one putative glycosylation site, while Gc has a larger ectodomain (~900 amino acids) predicted to be a class II fusion protein with 3-4 glycosylation sites.11 These glycoproteins form heterodimers in the endoplasmic reticulum lumen and are transported to Golgi apparatus for virion assembly, ultimately creating distinctive trimeric spike structures on the viral surface that mediate host cell attachment and entry.22,23 The NSm protein, though its exact function remains unclear, may participate in viral assembly and budding but appears dispensable for replication based on reverse genetics studies.24 The S segment (~946 nucleotides) encodes for both the nucleocapsid protein primarily forms the capsid and stabilizes the genome and the non-structural NSs protein translated from an alternative start site on the same mRNA. While not incorporated into virions, NSs plays critical roles in viral replication and immune evasion by suppressing host interferon responses, particularly through its C-terminal nine amino acids in OROV (Figure 2).24
The viral replication process occurs in the cytoplasm of the infected cell. In this process, the RNA-dependent RNA polymerase enzyme is responsible for viral replication. This enzyme catalyzes the synthesis of viral genomic RNA. These newly synthesized RNA strands are then encapsidated by N proteins to form nucleocapsids. These nucleocapsids then move to the cell membrane and bind to the Gn and Gc proteins. This process is facilitated by lipid rafts, which are membrane microdomains containing sphingolipids and cholesterol. These lipid rafts are considered an essential platform for viral organization.11,25
The OROV strategically co-opts the host cell’s endosomal sorting complex essential for transport (ESCRT) machinery to execute critical membrane remodelling events during the budding process. Immunoprecipitation and fluorescence microscopy studies revealed specific recruitment of ESCRT components particularly charged multivesicular body protein 6 (CHMP6) to Golgi membranes upon OROV glycoprotein expression.26 This represents a novel adaptation of the conventional ESCRT-mediated budding pathway, which typically functions at endosomal membranes. Furthermore, the virus exploits the host vesicular transport system by hijacking Rab27a, a small GTPase that regulates membrane trafficking. The OROV controls Rab27a-dependent pathways to facilitate both intracellular transport of viral components and subsequent release of mature virions from infected cells. This dual exploitation of ESCRT machinery and Rab GTPase networks underscores the sophisticated mechanisms OROV employs to ensure efficient viral egress.27
Structural studies of related Oropouche bunyavirus particles (OBVs) like Bunyamwera virus reveal these surface glycoprotein spikes are formed by N-terminal extensions of Gc and represent primary targets for neutralizing antibodies, with immunization of mice against these domains inducing near-sterilizing immunity.22,23 Potential reassortment processes are facilitated by this genomic organization and structural architecture, which also determines critical elements of viral pathogenicity, host interactions, and antigenic characteristics crucial for vaccine development and diagnostics.
Reassortment and evolutionary analysis
The segmented genome of OROV facilitates RNA segments from several viral strains to exchange during co-infection of a host cell in a process known as reassortment. When multiple viral strains co-infect a host, the segmented structure of OROV’s tripartite genome allows for possible reassortment processes giving rise to new viral strains. Novel viral variants with unique genomic profiles with altered host range, improved immune evasion skills, changed transmission dynamics and increased virulence can be produced by such genetic recombination. Emerging research has documented the appearance of new reassorted OROV strains that have been implicated in recent regional outbreaks. These evolved variants demonstrate concerning clinical characteristics, including increased pathogenicity, capacity for vertical transmission resulting into maternal-fetal infections, congenital abnormalities and increased transmission efficiency within human populations.28,29
Phylogenetic analysis of OROV strains from different regions reveals significant genetic diversity, particularly in the S segment. For example, strains from Ecuador and Brazil show high homology in the L and M segments but differ in the S segment, suggesting reassortment events.11,30 Related viruses have been seen to reassort, including the Ngari virus which is a reassortant between the Batai and Bunyamwera viruses that causes severe haemorrhagic fever in humans.31 Similarly, Madre de Dios virus in Venezuela and Iquitos virus (IQTV) in Peru are thought to have emerged through reassortment events.32,33 These findings highlight the potential for OROV to evolve into a more pathogenic version of itself and cause more severe disease.
Transmission
The OROV is naturally maintained in two distinct cycles viz. urban cycle and sylvatic cycle (Figure 3). The urban cycle involves Culicoides paraensis biting midges, which feed on blood of humans and transmit the virus into the blood stream simultaneously.34 These biting midges serve as competent vectors not only for OROV but also for several other medically important arboviruses including Akabane virus (AKAV), bluetongue virus (BTV), bovine ephemeral fever virus (BEFV), epizootic hemorrhagic disease virus (EHDV), and African horse sickness virus (AHSV).35,36 The genus Culicoides comprises over 1,300 species worldwide, with approximately 13% remaining unclassified, and exhibits a broad geographic distribution ranging from Argentina to different parts of the northern United States for C. Paraensis.37 Although C. paraensis, has distinct diurnal activity peaks and behavioural plasticity by biting both indoors and outdoors in shaded areas, with increased activity during rainy seasons and decreased activity during hot, dry ones, adult Culicoides are primarily crepuscular in their biting habits.6 It has been hypothesized the virus can also be transmitted by species of mosquitoes such as Culex quinquefasciatus and Aedes aegypti, though C. paraensis remains the primary vector.25,38,39 Experimental studies have confirmed that C. paraensis can effectively transmit OROV biologically, demonstrating an 83% transmission rate between infected and susceptible hamsters, as well as between infected humans and susceptible hamsters.40
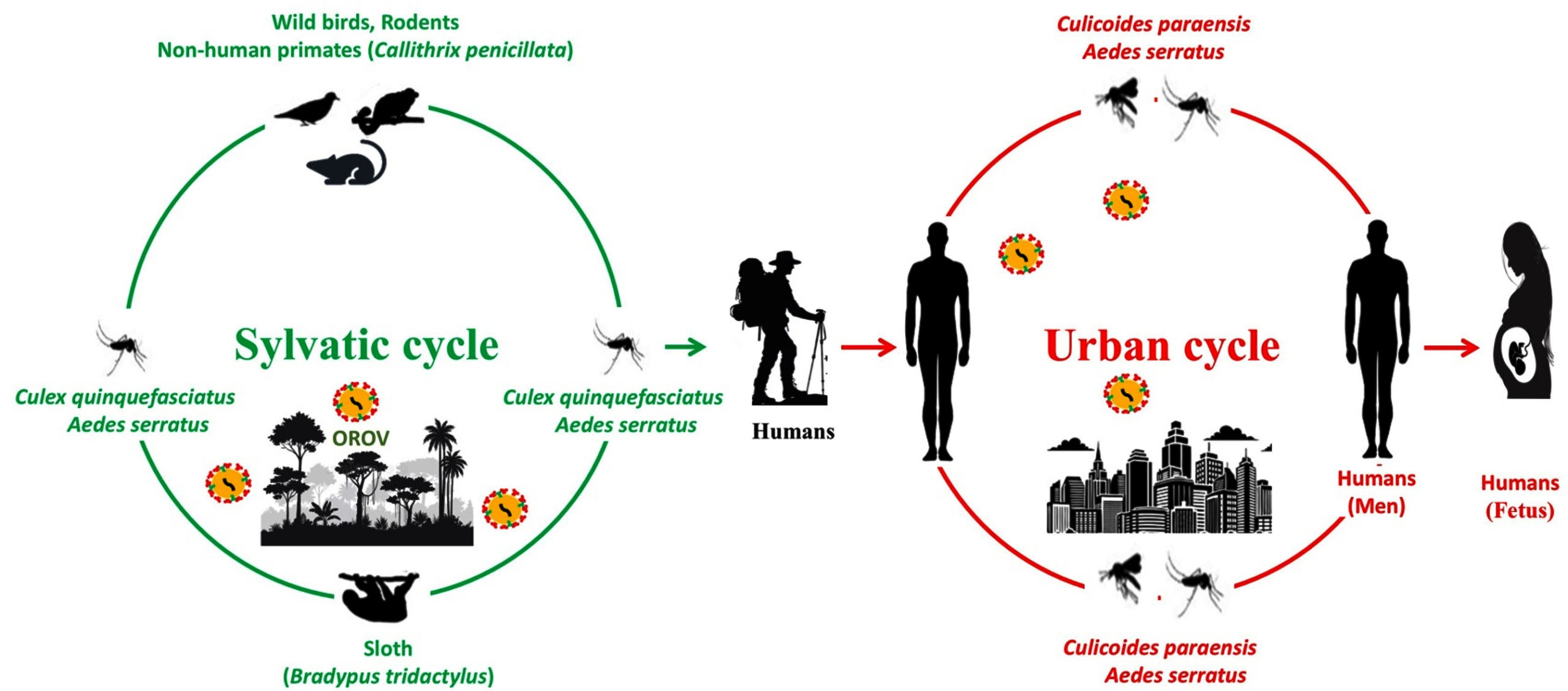
Figure 3. Schematic representation of the dual-cycle (sylvatic and urban) transmission of Oropouche virus, illustrating the intersection between animal reservoirs, insect vectors and the spillover into human populations
The sylvatic cycle involves wild animals such as sloths (Bradypus tridactylus) and non-human primates, which serve as reservoir hosts. The fundamental function of the sylvatic cycle is zoonotic maintenance among wild animal reservoirs, which include non-human primates, some avian hosts, and several mammalian species, especially sloths.41 Although serological evidence points to possible viral circulation in these wildlife groups, more research is necessary to confirm their reservoir competence.
Anthropogenic environmental changes, such as climate, deforestation, mining, agricultural activities and expansion of urban areas coupled with the adaptability of Culicoides species aid in the shift from sylvatic to urban transmission resulting in to increase in the risk of OROV emergence in new areas. By increasing human-vector contact, these ecological disruptions have the potential to connect the two cycles of transmission. Notably, there are no verified instances of domestic animals being involved in viral amplification or transmission, making humans the only vertebrate hosts in the urban transmission cycle.42 While some studies have hypothesized potential amplification roles for domestic poultry, empirical evidence supporting this hypothesis remains lacking.
Vertical transmission
In addition to arthropod-borne transmission, OROV exhibits the ability to spread vertically.43 Recent studies have demonstrated that the OROV can cross the placental barrier and lead to congenital infection and severe adverse pregnancy complications.44 These include miscarriage, stillbirth, and congenital abnormalities, particularly those affecting the central nervous system (CNS), such as microcephaly.45 This pathogenic feature is similar to that of other Simbu serogroup orthobunyaviruses that infect ruminants congenitally.46 The identification of replication-competent OROV in semen raises the possibility of transmission risks comparable to those described for the Zika virus, despite the fact that no cases of transfusion-mediated or sexual transmission have been observed.47,48 However, the epidemiological significance of such potential transmission routes remains unconfirmed.
As of December 2024, Brazil has reported three laboratory-confirmed cases of vertical transmission leading to two stillbirths and one congenital defect. Additionally, investigation of 15 fetal deaths, four cases of miscarriages and five cases of congenital anomalies potentially linked to OROV infection during pregnancy are still being carried out.49 These findings highlight the virus’s potential to cause significant perinatal morbidity and similar to other arboviruses (e.g. Zika), its tropism for the placenta and foetus, raising concerns about its impact on pregnancy. These observations reinforce the importance of prenatal screening protocols in endemic regions and advising pregnant individuals on preventive measures against vector-borne transmission.
Clinical disease
Following an incubation period of typically three to eight days, clinical symptoms like fever, muscle pain, headache and joint pain appear. In extreme illness, the virus may result in neurological problems like meningitis and encephalitis or haemorrhagic symptoms including epistaxis and gingival bleeding.50 Recurrence of symptoms is common, with approximately 60% of patients experiencing a second wave of fever and other symptoms one to two weeks after recovery.25,51 Additionally, cases of pregnancy complications, including microcephaly, congenital abnormalities and fetal death has also been reported.52 Although the mechanisms causing recurrent febrile sickness are unknown, they can include an insufficient immune response or viral persistence in immune-privileged areas (Figure 4).
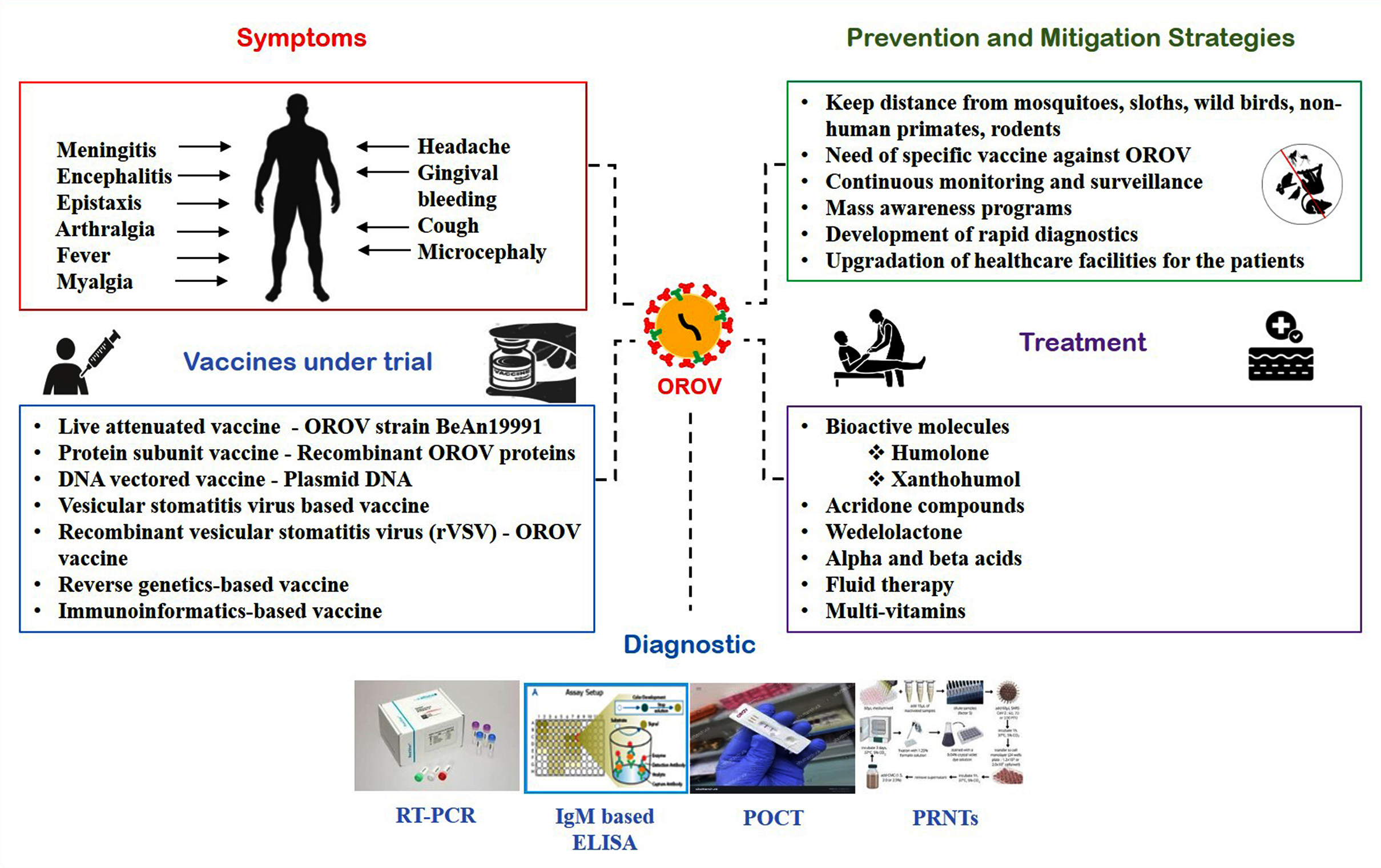
Figure 4. Illustration of the clinical signs of Oropouche virus detailing the progression of symptomatic manifestations alongside current public health interventions. The illustration categorizes management into three pillars: evidence-based prevention and mitigation techniques, current supportive treatment protocols, and the latest advancements in experimental vaccine candidates currently undergoing clinical and preclinical trials
Pathogenesis
When pattern-recognition receptors (PRRs) identify viral pathogen-associated molecular patterns (PAMPs) during viral infections, the host immune system is immediately activated. Toll-like receptors (TLRs) and RIG-I like receptors (RLRs) detects the OROV genome as a crucial PAMP, which triggers the release of inflammatory and antiviral cytokines. Research utilizing murine models has shown that the innate immune response to OROV is facilitated by the type I IFN pathway, which includes interferon regulatory factors (IRF3, IRF5, and IRF7), IFN-α/β receptors, and the mitochondrial antiviral signalling protein (MAVS).53 The IRF5 has been identified as a crucial regulator of type I IFN responses in peripheral tissues, governing OROV and the related neurotropic virus La Crosse virus (LACV), thus restricting neuroinvasion. Further investigations using OROV and lymphocytic choriomeningitis virus (LCMV) infection models revealed that MAVS, IRF3, IRF7, and IFNAR restrict viral replication and mitigate liver damage.54 Bone marrow chimera experiments highlighted the importance of IFN signalling in non-myeloid cells, as Ifnar-/- mice reconstituted with wild-type hematopoietic cells still exhibited high viral loads and liver injury, whereas wild-type mice with Ifnar-/- bone marrow remained resistant. Additionally, IFN-β-/- mice showed partial susceptibility (17% lethality), while Ifnar-/- mice suffered complete lethality, underscoring the essential role of IFN-α in controlling OROV infection. In vitro studies indicated a transient up regulation of IFN-β mRNA within the first hour of infection, followed by a sharp decline by 24 hours. Type III IFN levels also increased significantly, contributing to antiviral defence; whereas type II IFN (IFN‐γ) peaked in peripheral blood mononuclear cells (PBMCs) at 24 hours before slightly decreasing by 48 hours.54 Low serum IFN‐γ levels in OROV fever patients suggest its immunomodulatory role in disease progression.55
The neurotropic potential of OROV has been confirmed by its isolation from the cerebrospinal fluid (CSF) of infected patients. Subcutaneous OROV injection caused CNS invasion in neonatal BALB/c mice, and the virus propagated from the periaqueductal gray to the forebrain. Widespread brain infection and severe encephalitis result from early infection of neural pathways, most likely through retrograde axonal transport, followed by penetration of the blood-brain barrier.56 The OROV can also persist at low levels in human immune cells (T cells, monocytes, dendritic cells, and B cells), suggesting a potential “Trojan horse” mechanism for entry into CNS to get beyond the blood-brain barrier.57 After entering the central nervous system, the virus can multiply in neurons, resulting in neurological disorders such as encephalitis and meningitis. Once inside the CNS, OROV replicates in neurons, causing inflammation and potentially leading to long-term neurological deficits. Mice experiments have shown that OROV infection can result in severe encephalitis, with glial cells and neurons hosting viral replication.58,59
Immunity against infection
The host defence mechanism for OROV infection is governed by a two-part immunological principle that unites innate immunity with specific immunity. The primary step of OROV infection triggers an instant innate immunity response, which is primarily exemplified by the production of type I interferons (IFN) coupled with a subsequent cascade of pro-inflammatory cytokines, which are intended to control early viral titers. Despite this, OROV has developed complex immune evasive phenotypes. The virus can evade the host immune system through modulation of cytokine signaling pathways, leading to an impairment of the functional capacity of key immune cells.55,60
Diagnostic methods
Oropouche virus induced fever frequently goes undiagnosed due to its mild clinical presentation and overlapping symptoms with other arboviral infections like dengue, Zika, chikungunya, malaria, leptospirosis or yellow fever.38 Routine haematological analyses including complete blood counts (CBC) have restricted diagnostic accuracy, and findings like leukopenia and elevated alanine transaminase (ALT) levels are inconclusive and occur in other febrile illnesses. Examination of CSF in neuroinvasive cases may reveal pleocytosis, altered glucose levels, and elevated protein concentrations, which are commonly observed in various viral encephalitides, are not typical for OROV.61 Serological interpretation is complicated by potential cross-reactivity with antibodies against related arboviruses, necessitating confirmatory testing. Furthermore, it is imperative to develop cell lines and animal models to study OROV pathogenesis and host immune responses. Although virus isolation in permissive cell cultures is practicable, its value is restricted by protracted downtime and capital-intensive protocols.62 The primary diagnostic methods include:
Detection of OROV nucleic acid
Clinical samples might contain OROV RNA, which can be identified using molecular techniques like reverse transcription-polymerase chain reaction (RT-PCR). The RT-PCR assays focusing on M or S genomic segments have become the most dependable means of identifying OROV RNA in clinical specimens (e.g., blood, saliva, and urine) during the acute phase.35,63-65 Although having a high specificity, the sensitivity of RT-PCR assays can range depending on viral load and sample quality. Early virus detection during the acute stage of infection is made possible by this highly specific approach.
Detection of OROV immunoglobulin M (IgM)
Serological testing is fundamental to diagnosing OROV. Enzyme-linked immunosorbent assay (ELISA) tests are used to identify IgM antibodies, which are produced early in the immunological response. Serum or CSF containing OROV-specific IgM indicates a recent acute OROV infection.
Detection of OROV neutralizing antibodies
In immunocompetent patients OROV-specific neutralizing antibodies are identifiable through plaque reduction neutralization tests (PRNTs), and IgM seroconversion usually takes place within the initial week after symptoms begin. A conclusive diagnosis often requires exhibiting a four-fold rise in neutralizing antibody titers between paired acute and convalescent serum samples.49 When molecular testing is either unavailable or inconclusive, this technique is especially helpful for verifying OROV infection. Because clinical signs of OROV are often non-specific and resembles with other febrile diseases, it is essential to train healthcare professionals in distinguishing OROV from other clinically similar arboviral diseases to enhance diagnostic accuracy.
At present, confirmatory testing for OROV depends on specialized serological assays that are not commercially accessible and the tests are limited to reference laboratories such as the United States Centers for Disease Control and usually conducted in collaboration with state or territorial health departments (Figure 4). This limitation highlights the necessity for greater accessibility to validated diagnostic tools in endemic regions, in order to enable timely and precise detection of OROV infections.66
Vaccine development
Considering the pandemic potential of the OROV, production of an OROV vaccine must be of prime concern for global researchers and health agencies (Figure 4). Several vaccine candidates are under investigation including inactivated, live attenuated and recombinant protein-based vaccines. A live attenuated vaccine based on the Belem, Ananindeua (BeAn19991) strain has shown promising results in preclinical studies, inducing robust immune responses in animal models.11,67 Reverse genetics systems are being investigated for the development of vaccines that generate robust and specific immune response.68 One promising candidate for vaccine development is a replicable vesicular stomatitis virus (VSV) expressing OROV glycoproteins which can elicit an immune response in the host body. This vaccine has been shown to protect mice from wild-type OROV challenge.67 Additionally, Immunoinformatics approaches have identified potential B and T cell epitopes within the M-segment polyprotein of OROV, offering a crucial strategy for vaccine development.26
Treatment
At present, no approved antiviral treatments are available for OROV infection. Significant broad-spectrum antiviral action has been demonstrated by acridone derivatives against a variety of viruses such as RNA viruses from the Flaviviridae and other DNA viruses from the Herpesviridae family (Figure 4). Specifically, acridone compounds FAC21 and FAC22 have demonstrated potent inhibition of OROV replication in vitro, achieving a 99.9% reduction in viral yield through targeted interaction with the viral endonuclease.69 This enzymatic target plays a critical role in the cap-snatching mechanism characteristic of orthobunyaviruses, representing a promising aspect for antiviral intervention. Complementary studies have identified natural compounds with anti-OROV activity, including hop (Humulus lupulus L.) derived constituents such as humulone, alpha acids, beta acids, xanthohumol and flavonoids. These bioactive molecules effectively suppressed OROV replication during the early stages of infection (12-48 hours post-infection) by interfering with endonuclease function.70 Furthermore, computational and experimental analyses have highlighted Wedelolactone (WDL) as a potential inhibitor of OROV endonuclease, demonstrating in silico binding affinity and subsequent in vitro antiviral effects across multiple stages of viral infection in mammalian cell models.71 These findings collectively underscore the therapeutic potential of targeting the OROV endonuclease for antiviral drug development.
Prevention and control
In absence of effective antiviral treatment for OROV infection, preventive measures become paramount (Figure 4). Furthermore, the absence of a licensed vaccine complicates the efforts made in order to control the OROV. Management strategies depend on a threefold set of interventions: (1) Protective measures aimed at individuals to minimize skin exposure like wearing long-sleeved clothes and long pants and installing window and door screens to prevent vector entry into houses. (2) Vector control initiatives designed to restrict transmission. Reducing exposure to biting midges and mosquitoes is the mainstay of preventive interventions. Strategies for preventing the disease comprises of utilizing insect repellent with N,N-diethyl-meta-toluamide (DEET) or picradin as their active ingredient. Eliminating standing water sources, such as containers and puddles, where biting midges and mosquitoes breed will also obviate the disease. (3) Supportive treatment for clinical symptoms.25 Management of the disease focuses on supportive care, including hydration, pain relief, and monitoring for complications such as neurological involvement. In severe cases, hospitalization may be required to manage symptoms and prevent long-term sequelae. Efficient outbreak management can be achieved with a decrease in the public health impact of OROV infections if these measures are strictly taken.
Oropouche virus poses a significant public health risk in South America with the high potential to spread to new regions due to environmental and climatic changes. The designing of a vaccine is complicated by the virus’s segmented genome and reassortment capabilities. However, advances in immunoinformatics, reverse genetics, and vaccine platforms offer promising strategies for preventing OROV transmission. Continued research and international collaboration are essential to develop effective vaccines and control measures, ultimately reducing the burden of Oropouche fever worldwide. This review provides a detailed explanation of OROV neuroinvasion, diagnostic methods, treatment options, and preventive strategies. It emphasizes the role of healthcare providers in identifying and reporting cases as well as the implementation of public health measures such as vaccine development for proper containment of the OROV.
ACKNOWLEDGMENTS
The authors would like to express their sincere gratitude to their respective institutions for their invaluable support during the preparation of this comprehensive review.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
AA conceptualized the study. AA, PPL, AG, SKP, and RG performed the literature review and data curation. AK, K, KG, and AKS carried out data visualization and designed the figures. AA, PPL, SP, and AKN wrote the manuscript. KCN, JR, and MS reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Watts DM, Phillips I, Callahan JD, Griebenow W, Hyams KC, Hayes CG. Oropouche virus transmission in the Amazon River basin of Peru. Am J Trop Med Hyg. 1997;56(2):148-152.
Crossref - Baisley KJ, Watts DM, Munstermann LE, Wilson ML. Epidemiology of endemic Oropouche virus transmission in upper Amazonian Peru. Am J Trop Med Hyg. 1998;59(5):710-716.
Crossref - Culquichicon C, Cardona-Ospina JA, Patino-Barbosa AM, Rodriguez-Morales AJ. Bibliometric analysis of Oropouche research: impact on the surveillance of emerging arboviruses in Latin America. F1000Res. 2017;6:194.
Crossref - Acrani GO, Tilston-Lunel NL, Spiegel M, et al. Establishment of a minigenome system for Oropouche virus reveals the S genome segment to be significantly longer than reported previously. J Gen Virol. 2015;96(3):513-523.
Crossref - de Souza LAF, Sena da Silva SL, do Nascimento BLS, de Sousa Farias E, Nunes Neto JP, Costa-Ribeiro MCVD. First report of Culicoides paraensis (Goeldi, 1905) (Diptera: Ceratopogonidae) in the state of Parana, Southern Brazil. Acta Trop. 2025;271:107868.
Crossref - Gallichotte EN, Ebel GD, Carlson CJ. Vector competence for Oropouche virus: A systematic review of pre-2024 experiments. PLoS Negl Trop Dis. 2025;19(4):e0013014.
Crossref - Mourao MP, Bastos MS, Gimaque JB, et al. Oropouche fever outbreak, Manaus, Brazil, 2007-2008. Emerg Infect Dis. 2009;15(12):2063-2064.
Crossref - de Souza Bastos M, Figueiredo LT, Naveca FG, et al. Identification of Oropouche Orthobunyavirus in the cerebrospinal fluid of three patients in the Amazonas, Brazil. Am J Trop Med Hyg. 2012;86(4):732-735.
Crossref - Pastula DM, Beckham JD, Tyler KL. Oropouche virus: an emerging neuroinvasive arbovirus. Ann Neurol. 2024:97(1):28-33.
Crossref - Gourjault C, Pezzi L, Doudier B, et al. Persistence of Oropouche virus in body fluids among imported cases in France, 2024. Lancet Infect Dis. 2025;25(2):e64-e65.
Crossref - Files MA, Hansen CA, Herrera VC, et al. Baseline mapping of Oropouche virology, epidemiology, therapeutics, and vaccine research and development. NPJ Vaccines. 2022;7(1):38.
Crossref - Cardoso BF, Serra OP, Heinen LBD, et al. Detection of Oropouche virus segment S in patients and in Culex quinquefasciatus in the state of Mato Grosso, Brazil. Mem Inst Oswaldo Cruz. 2015;110(6):745-754.
Crossref - Travassos da Rosa JF, de Souza WM, Pinheiro FP, et al. Oropouche virus: clinical, epidemiological, and molecular aspects of a neglected orthobunyavirus. Am J Trop Med Hyg. 2017;96(5):1019-1030.
Crossref - World Health Organization. Oropouche Virus Disease—Region of the Americas. 2024. https://www.who.int/emergencies/disease -outbreak-news/item/2024-DON530. Accessed February 17, 2025.
- Deiana M, Malago S, Mori A, et al. Full genome characterization of the first Oropouche virus isolate imported in Europe from Cuba. Viruses. 2024;16(10):1586.
Crossref - Morrison A, White JL, Hughes HR, et al. Oropouche virus disease among U.S. travelers — United States, 2024. MMWR Morb Mortal Wkly Rep. 2024;73(35):769-773.
Crossref - Centers for Disease Control and Prevention. CDC Oropouche current year data 2025. https://www.cdc.gov/oropouche/data-maps/current-year-data.html. Accessed February 17, 2025.
- Pan American Health Organization. Epidemiological update: Oropouche in the region of the Americas. 2025. https://www.paho. org/en/documents/epidemiological- update-oropouche-region-americas- 13-august-2025.Accessed February 17, 2025.
- Gaillet M, Pichard C, Restrepo J, et al. Outbreak of Oropouche virus in French Guiana. Emerg Infect Dis. 2021;27(10):2711-2714.
Crossref - Ciuoderis KA, Berg MG, Perez LJ, et al. Oropouche virus as an emerging cause of acute febrile illness in Colombia. Emerg Microbes Infect. 2022;11(1):2645-2657.
Crossref - Tilston-Lunel NL, Hughes J, Acrani GO, et al. Genetic analysis of members of the species Oropouche virus and identification of a novel M segment sequence. J Gen Virol. 2015;96(7):1636-1650.
Crossref - Bowden TA, Bitto D, McLees A, Yeromonahos C, Elliott RM, Huiskonen JT. Orthobunyavirus ultrastructure and the curious tripodal glycoprotein spike. PLoS Pathog. 2013;9(5):e1003374.
Crossref - Hellert J, Aebischer A, Wernike K, et al. Orthobunyavirus spike architecture and recognition by neutralizing antibodies. Nat Commun. 2019;10(1):879.
Crossref - Tilston-Lunel NL, Acrani GO, Randall RE, Elliott RM. Generation of recombinant Oropouche viruses lacking the nonstructural protein NSm or NSs. J Virol. 2016;90(5):2616-2627.
Crossref - Pan American Health Organization. Epidemiological update: Oropouche in the region of the Americas. 2025. https://www.paho.org/en/documents/epidemiological-update-oropouche-region-americas-13-august-2025 Accessed February 17, 2025
- Barbosa NS, Concha JO, Dasilva LLP, Crump CM, Graham SC. Oropouche virus glycoprotein topology and cellular requirements for glycoprotein secretion. J Virol. 2023;97(1):e0133122.
Crossref - Concha JO, Gutierrez K, Barbosa N, et al. Rab27a GTPase and its effector Myosin Va are host factors required for efficient Oropouche virus cell egress. PLoS Pathog. 2024;20(8):e1012504.
Crossref - Naveca FG, de Almeida TAP, Souza V, et al. Human outbreaks of a novel reassortant Oropouche virus in the Brazilian Amazon region. Nat Med. 2024;30(12):3509-3521.
Crossref - Schwartz DA. Novel reassortants of Oropouche virus (OROV) are causing maternal-fetal infection during pregnancy, stillbirth, congenital microcephaly and malformation syndromes. Genes. 2025;16(1):87.
Crossref - Gutierrez B, Wise EL, Pullan ST, et al. Evolutionary dynamics of Oropouche virus in South America. J Virol. 2020;94(9):e00121-20.
Crossref - Briese T, Bird B, Kapoor V, Nichol ST, Lipkin WI. Batai and Ngari viruses: M segment reassortment and association with severe febrile disease outbreaks in East Africa. J Virol. 2006;80(11):5627-5630.
Crossref - Aguilar PV, Barrett AD, Saeed MF, et al. Iquitos virus: a novel reassortant Orthobunyavirus associated with human illness in Peru. PLoS Negl Trop Dis. 2011;5(9):e1315.
Crossref - Navarro JC, Giambalvo D, Hernandez R, et al. Isolation of Madre de Dios virus (Orthobunyavirus; Bunyaviridae), an Oropouche virus species reassortant, from a monkey in Venezuela. Am J Trop Med Hyg. 2016;95(2):328-338.
Crossref - Centers for Disease Control and Prevention. Clinical overview of Oropouche virus disease. Published 2025. https://www.cdc.gov/oropouche/hcp/clinical-overview/index.html. Accessed January 28, 2025.
- Sick F, Beer M, Kampen H, Wernike K. Culicoides biting midges – underestimated vectors for arboviruses of public health and veterinary importance. Viruses. 2019;11(4):376.
Crossref - Saminathan M, Singh KP, Khorajiya JH, et al. An updated review on bluetongue virus: epidemiology, pathobiology, and advances in diagnosis and control with special reference to India. Vet Q. 2020;40(1):258-321.
Crossref - Dunford JC, Vigil SL, Ruder MG, et al. A review and update of the distribution, bionomics, and medical importance of Culicoides (Haematomyidium) paraensis (Diptera: Ceratopogonidae) in the United States in response to recent Oropouche virus expansion in the Americas. J Med Entomol. 2026;63(1):tjag019.
Crossref - Pinheiro FP, Travassos da Rosa AP, Travassos da Rosa JF, et al. Oropouche virus. I. A review of clinical, epidemiological, and ecological findings. Am J Trop Med Hyg. 1981;30(1):149-160
- Nunes MRT, Martins LC, Rodrigues SG, et al. Oropouche virus isolation, southeast Brazil. Emerg Infect Dis. 2005;11(10):1610-1613.
Crossref - Bandeira AC, da Silva Barbosa ACFN, Souza M, et al. Clinical profile of Oropouche fever in Bahia, Brazil: unexpected fatal cases. SciELO Prepr. 2024.
Crossref - Sciancalepore S, Schneider MC, Kim J, Galan DI, Riviere-Cinnamond A. Presence and multi-species spatial distribution of Oropouche virus in Brazil within the One Health framework. Trop Med Infect Dis. 2022;7(6):111.
Crossref - da Rosa JF, de Souza WM, de Paula Pinheiro F, et al. Oropouche virus: clinical, epidemiological, and molecular aspects of a neglected orthobunyavirus. Am J Trop Med Hyg. 2017;96(5):1019-1030.
Crossref - Schwartz DA, Baud D, Dashraath P. A potential mechanism of transplacental transmission of Oropouche virus in pregnancy. Lancet Microbe. 2025;6(7):101083.
Crossref - Dashraath P, Nielsen-Saines K, Schwartz DA, Musso D, Baud D. Vertical transmission potential of Oropouche virus infection in human pregnancies. AJOG Glob Rep. 2025;5(1):100431.
Crossref - das Neves Martins FE, Chiang JO, Nunes BTD, et al. Newborns with microcephaly in Brazil and potential vertical transmission of Oropouche virus: a case series. Lancet Infect Dis. 2025;25(2):155-165.
Crossref - O’Connor TW, Hick PM, Finlaison DS, Kirkland PD, Toribio JAL. Revisiting the importance of orthobunyaviruses for animal health: a scoping review of livestock disease, diagnostic tests, and surveillance strategies for the Simbu serogroup. Viruses. 2024;16(2):294.
Crossref - Castilletti C, Huits R, Mantovani RP, et al. Replication-competent Oropouche virus in semen of traveler returning to Italy from Cuba, 2024. Emerg Infect Dis. 2024;30(12):2684-2686.
Crossref - Counotte MJ, Kim CR, Wang J, et al. Sexual transmission of Zika virus and other flaviviruses: a living systematic review. PLoS Med. 2018;15(7):e1002611.
Crossref - Pan American Health Organization. Epidemiological alert: Oropouche in the region of the Americas: vertical transmission event under investigation in Brazil. 2024. https://www.paho.org/en/documents/epidemiological-alert-oropouche-region-americas-vertical-transmission-event-under. Accessed September 18, 2024.
- Barrera ELP, Reales-Gonzalez J, Salas D, et al. Fatal acute undifferentiated febrile illness among clinically suspected leptospirosis cases in Colombia, 2016–2019. PLoS Negl Trop Dis. 2023;17(4):e0011683.
Crossref - Da Costa Vasconcelos PF, Da Rosa JFST, Guerreiro SC, Dégallier N, Da Rosa EST, De Andrade Travassos Da Rosa AP. Epidemic outbreaks of Oropouche virus in the Maranhão and Goiás States, Brazil. Revista Do Instituto De Medicina Tropical De São Paulo. 1989;31(4):271-278.
Crossref - Schwartz DA, Dashraath P, Baud D. Oropouche virus (OROV) in pregnancy: an emerging cause of placental and fetal infection associated with stillbirth and microcephaly following vertical transmission. Viruses. 2024;16(9):1435.
Crossref - Proenca-Modena JL, Hyde JL, Sesti-Costa R, et al. Interferon-regulatory factor 5-dependent signaling restricts orthobunyavirus dissemination to the central nervous system. J Virol. 2016;90(1):189-205.
Crossref - Proenca-Modena JL, Sesti-Costa R, Pinto AK, et al. Oropouche virus infection and pathogenesis are restricted by MAVS, IRF-3, IRF-7, and type I interferon signaling pathways in nonmyeloid cells. J Virol. 2015;89(9):4720-4737.
Crossref - de Oliveira E, Azevedo RD, Coelho-dos-Reis JG, et al. IFN-a as a time-sensitive biomarker during Oropouche virus infection in early and late seroconverters. Sci Rep. 2019;9(1):17924.
Crossref - Almeida GM, Souza JP, Mendes ND, et al. Neural infection by Oropouche virus in adult human brain slices induces an inflammatory and toxic response. Front Neurosci. 2021;15:674576.
Crossref - Amorim MR, Cornejo Pontelli M, Fabiano de Souza G, et al. Oropouche virus infects, persists and induces IFN response in human peripheral blood mononuclear cells as identified by RNA PrimeFlow and qRT-PCR assays. Viruses. 2020;12(7):785.
Crossref - Santos RI, Bueno-Junior LS, Ruggiero RN, et al. Spread of Oropouche virus into the central nervous system in mouse. Viruses. 2014;6(10):3827-3836.
Crossref - da Silva Menegatto MB, Ferraz AC, Lima RLS, et al. Oropouche virus infection induces ROS production and oxidative stress in liver and spleen of mice. J Gen Virol. 2023;104(5):1857.
Crossref - Elliott RM. Orthobunyaviruses: recent genetic and structural insights. Nat Rev Microbiol. 2014;12(10):673-685.
Crossref - Bastos MS, Lessa N, Naveca FG, et al. Detection of Herpesvirus, Enterovirus, and Arbovirus infection in patients with suspected central nervous system viral infection in the Western Brazilian Amazon. J Med Virol. 2014;86(9):1522-1527.
Crossref - Durango-Chavez HV, Toro-Huamanchumo CJ, Silva-Caso W, et al. Oropouche virus infection in patients with acute febrile syndrome: is a predictive model based solely on signs and symptoms useful? PLoS One. 2022;17(7):e0270294.
Crossref - Fonseca L, Carvalho RH, Bandeira AC, et al. Oropouche virus detection in febrile patients’ saliva and urine samples in Salvador, Bahia, Brazil. Jpn J Infect Dis. 2020;73(2):164-165.
Crossref - Nascimento VAD, Santos JHA, Monteiro DCDS, et al. Oropouche virus detection in saliva and urine. Mem Inst Oswaldo Cruz. 2020;115:e190338.
Crossref - Wesselmann KM, Postigo-Hidalgo I, Pezzi L, et al. Emergence of Oropouche fever in Latin America: a narrative review. Lancet Infect Dis. 2024;24(7):e439-e452.
Crossref - Ribas Freitas AR, Schwartz DA, Lima Neto AS, et al. Oropouche virus (OROV): expanding threats, shifting patterns, and the urgent need for collaborative research in Latin America. Viruses. 2025;17(3):353.
Crossref - Stubbs SH, Pontelli MC, Mishra N, et al. Vesicular stomatitis virus chimeras expressing the Oropouche virus glycoproteins elicit protective immune responses in mice. mBio. 2021;12(3):e00463-21.
Crossref - Adhikari UK, Tayebi M, Rahman MM. Immunoinformatics Approach for Epitope-Based Peptide Vaccine Design and Active Site Prediction against Polyprotein of Emerging Oropouche Virus. J Immunol Res. 2018;2018:6718083.
Crossref - Saivish MV, Menezes GL, da Silva RA, et al. Acridones as promising drug candidates against Oropouche virus. Curr Res Microb Sci. 2024;6:100217.
Crossref - Mandova T, Saivish MV, Menezes GL, et al. Antiviral activity and molecular dynamics simulation of hops compounds against Oropouche virus (Peribunyaviridae). Pharmaceutics. 2023;15(12):2769.
Crossref - Peinado RS, Saivish MV, Menezes GL, et al. The search for an antiviral lead molecule to combat the neglected emerging Oropouche virus. Curr Res Microb Sci. 2024;6:100238.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.