


ISSN: 0973-7510
E-ISSN: 2581-690X
Methicillin-resistance in Staphylococcus aureus is frequently associated with enhanced biofilm formation and the presence of biofilm-associated genes that contribute to treatment failure and persistence. This study characterized the phenotypic biofilm formation and genotypic profiles of 10 clinical methicillin-resistant Staphylococcus aureus (MRSA) isolates and evaluated the dose-dependent antibiofilm activity of the ethanolic extract of oregano (Plectranthus amboinicus). All isolates carried mecA, confirming methicillin resistance in all isolates. The biofilm-associated genes icaA, icaD, and sarA were detected, whereas bap was absent. Despite the absence of bap, all isolates exhibited moderate to strong biofilm formation, consistent with ica-dependent polysaccharide-mediated adhesion. Based on PCR-based detection of the agrII gene (presence/absence), the isolates were classified into agrII-positive (n = 5) and agrII-negative (n = 5) groups. Biofilm formation was predominantly moderate (0.140 < OD < 0.280), with two isolates showing strong biofilm formation (OD > 0.280). Extract dilutions (100%-3.125% v/v) were tested using a modified crystal violet assay. The agrII-positive isolates exhibited significantly lower biofilm inhibition (12.9% ± 7.4%) than agrII-negative isolates (33.0% ± 1.2%) at 100% v/v (P = 0.003). Nonlinear regression using a four-parameter log-logistic model generated distinct dose-response curves, with 95% confidence bands exhibiting minimal overlap (12.5%-100%). These findings suggest a potential association between agrII status and differential biofilm susceptibility to P. amboinicus extract. However, given the limited sample size and in vitro design, these results should be interpreted with caution. Overall, this study highlights the potential of P. amboinicus as an antibiofilm agent and supports further investigation of genotype-informed strategies against MRSA.
Methicillin-resistant Staphylococcus aureus, Biofilm-associated Genes, Oregano Plant Extract, Antibiofilm Activity
Methicillin-resistant Staphylococcus aureus (MRSA) remains a clinically significant pathogen associated with persistent and difficult-to-treat infections. While its global burden is well recognized, treatment failure is increasingly attributed not only to antimicrobial resistance but also to the ability of MRSA to form biofilms, which enhance survival under hostile conditions and reduce the efficacy of antibiotics.1,2
Biofilm formation plays a central role in MRSA persistence by protecting bacterial cells from host immune responses and limiting antibiotic penetration.3,4 These structured communities contribute to chronic and device-associated infections, for which conventional therapeutic strategies are often insufficient. Consequently, increasing attention has been directed toward understanding the molecular mechanisms underlying biofilm development, particularly the regulatory systems governing adhesion and maturation. Disruption of quorum sensing (QS)-regulated genes, such as the accessory gene regulator (agr) system, which governs intercellular signaling and biofilm maturation, may reduce bacterial adherence and virulence.5,6
Biofilm-associated genes, including icaA, icaD, sarA, and bap, are traditionally implicated in biofilm matrix formation; however, MRSA can also form biofilms through alternative ica-independent mechanisms, such as protein-mediated adhesion and quorum-sensing regulation. Among these, the agr system is a key modulator of virulence and biofilm dynamics, influencing both structural integrity and dispersal. Plant-based QS inhibitors show promise for attenuating biofilm formation; however, studies on readily available medicinal species remain limited.7 In particular, there is limited evidence on whether antibiofilm activity varies according to MRSA genotypic differences, especially in relation to quorum-sensing systems.
Oregano (Plectranthus amboinicus), although not among the ten medicinal plants officially endorsed by the Philippine Department of Health, is widely used in folk medicine and exhibits antibacterial properties.8 Its ethanolic extract contains phenolic compounds, such as carvacrol and thymol, which disrupt bacterial membranes, alter signaling pathways, and degrade biofilm matrix components.9-11
Despite these known properties, the relationship between MRSA genotype and the antibiofilm activity of P. amboinicus remains unclear. In this study, the phenotypic biofilm characteristics of clinically isolated MRSA were evaluated, along with genotypic profiling of biofilm-associated genes. The ethanolic extract of P. amboinicus was assessed for dose-dependent antibiofilm activity. We aimed to determine whether agrII status is associated with differential antibiofilm susceptibility, thereby addressing a gap in genotype-informed antibiofilm strategies.
Bacterial strains
Ten clinically isolated MRSA strains were obtained from a tertiary hospital Microbiology laboratory in Davao City. This study was conducted as a preliminary pilot investigation, and the sample size (n = 10) was selected to explore initial genotype–phenotype associations.
Isolates were collected from different patients, and duplicate isolates from the same patient were excluded. Inclusion criteria included confirmed Staphylococcus aureus isolates with methicillin resistance, while contaminated or mixed cultures were excluded. Clinical specimens were derived from diverse sources, including tracheal aspirates, wound and tissue samples, blood, urine, catheter tips, and abscesses.
Isolates were phenotypically identified by standard biochemical tests and confirmed using the automated VITEK system (bioMérieux, Marcy-l’Etoile, France). Identification confidence values (probability scores ≥95%) were used for species-level confirmation.
S. aureus ATCC 25923 was used as a positive control for the biofilm formation assays. Isolates were transported under cold-chain conditions in nutrient agar slants and subcultured on blood agar at 37 °C for 24 h. Colonies with round, convex morphology (1-4 mm diameter) and weak to clear β-hemolysis were confirmed on Mueller–Hinton agar supplemented with 4% NaCl and 6 µg/mL of oxacillin.
Assessment of the biofilm formation capability of the clinically isolated MRSA
Biofilm formation was quantified using a crystal violet microtiter plate assay.3,12 All experiments were performed using three biological replicates (independent cultures) and three technical replicates per condition. MRSA strains were cultured on blood agar at 37 °C for 24 hrs, and three colonies were inoculated into 5 mL tryptic soy broth (TSB) and incubated for 18 hrs. Bacterial suspensions were standardized to OD580 = 0.50-0.63 McFarland Standard using VITEK® DensiCHEK (bioMérieux, Marcy-l’Etoile, France) in phosphate-buffered saline (PBS; pH 7.4). A 100 µL bacterial suspension was added to 100 µL of TSB in 96-well plates and incubated for 24 hrs at 37 °C. Wells were washed twice with sterile PBS, fixed with 96% ethanol (15 min), stained with 2% crystal violet (5 min), and destained with 33% acetic acid. Absorbance was measured at 492 nm. Biofilm production was categorized as none, weak (0.070 < OD < 0.140), moderate (0.140 < OD < 0.280), or strong (0.280 < OD) based on the cut-off optical density (ODc = mean OD of blank + 3 × SD) as previously described.13
Screening the clinically isolated MRSA for biofilm-associated genes
Isolates demonstrating biofilm formation (weak to strong) were screened for biofilm-associated genes (icaA, icaD, bap, sarA, and agrII) using polymerase chain reaction (PCR). Genomic DNA was extracted using a GF-1 Bacterial DNA Extraction Kit (Vivantis Technologies, Malaysia; Cat. No. GF-BA-050). A 25 µL PCR mixture contained 2 µL template DNA, 1 µL of each primer, 12.5 µL 2× Taq Master Mix (Vivantis Technologies, Malaysia; Cat. No. PLMM01), and 8.5 µL of nuclease-free water. Amplification was performed in a miniPCR mini16X thermocycler (miniPCR Bio, Cambridge, MA, USA) under the following conditions: 30 cycles at 94 °C for 30 sec (denaturation), primer-specific annealing temperatures (40-50 °C depending on the primer set; exact temperatures are provided in Table 1) for 30 sec, and extension at 72 °C for 17-59 sec. Each PCR run included a negative control (no template control) and a reference strain (Staphylococcus aureus ATCC 25923) used as a control based on documented gene characteristics to verify amplification. PCR products were visualized by 1.5% agarose gel electrophoresis using the GELATOTM system (miniPCR Bio, Cambridge, MA, USA) at a running voltage of 135 V for 20-25 min. Primers were selected from a previously published study14 and validated for specificity, with the sequences and corresponding details presented in Table 1.
Table (1): Primer sequences and PCR conditions for the screening of biofilm-associated genes in clinically isolated MRSA
| Genes | Primer Sequence | Product size (bp) | Annealing time (s) |
|---|---|---|---|
| mecA | 5′-ATCGATGGTAAAGGTTGG-3′ | 533 | 32 |
| 5′-AGTTCTGCAGTACCGGATTTG-3′ | |||
| icaA | 5′-TGGCTGTATTAAGCGAAGTC-3′ | 669 | 41 |
| 5′-CCTCTGTCTGGGCTTGACC-3′ | |||
| icaD | 5′-AAACGTAAGAGAGGTGG-3′ | 381 | 23 |
| 5′-GGCAATATGATCAAGATAC-3′ | |||
| bap | 5′-CCCTATATCGAAGGTGTAGAATTG-3′ | 971 | 59 |
| 5′-GCTGTTGAAGTTAATACTGTACCTGC-3′ | |||
| sarA | 5′-TTAGCTTTGAAGAATTCGCTGT-3′ | 275 | 17 |
| 5′-TTCAATTTCGTTGTTTGCTTC-3′ | |||
| agrII | 5′-GTAGAGCCGTATTGATTCC-3” | 463 | 28 |
| 5′-GTATTTTCATCTCTTTAAGG-3 |
Plant extract preparation
Fresh P. amboinicus plants cultivated in Calinan, Davao city were authenticated by a botanist prior to experimentation. Mature leaves (2.5-3 cm long) were harvested before flowering, washed, and dehydrated at 30 °C to preserve phenolic compounds.15,16 Dried leaves were ground, and 5.5 g of powder was macerated in 100 mL of 60% ethanol for 7 days at ambient temperature with daily agitation. This process was repeated three times, and the extracts were pooled, filtered through Whatman No. 1 paper, and concentrated by rotary evaporation.17 The resulting extract exhibited an oily consistency and low recovery following lyophilization; therefore, the crude extract was used directly without determining the percentage yield. The crude extract was diluted in 5% dimethyl sulfoxide to prepare two-fold serial dilutions.18
Phytochemical Analysis
Qualitative phytochemical screening of the P. amboinicus ethanolic extract was performed using standard colorimetric assays. Phenols were detected using the ferric chloride test and characterized by the formation of a deep bluish-black coloration. Flavonoids were identified using the sodium hydroxide test, which produced an intense yellow color that turned colorless upon acidification. The presence of tannins was confirmed using the lead acetate test (yellowish precipitate), indicating their presence in the extract.
This analysis was limited to qualitative detection, and the quantitative determination of phytochemical constituents using chromatographic techniques such as high-performance liquid chromatography (HPLC) or gas chromatography–mass spectrometry (GC-MS) was not performed. As this study was designed as a preliminary pilot investigation, the phytochemical assessment aimed to provide an initial characterization of the extract. Therefore, the relative concentrations of the active compounds could not be determined.
Dose-response analysis of P. amboinicus ethanolic extract
The dose-dependent antibiofilm activity was evaluated using a modified crystal violet assay. Bacterial suspensions standardized to OD580 = 0.50-0.63 McFarland Standard using VITEK® DensiCHEK (bioMérieux, Marcy-l’Etoile, France) were co-incubated with equal volumes (50 µL) of serially diluted P. amboinicus ethanolic extract and 100 µL TSB in 96-well plates for 24 hrs at 37 °C. The experiments were conducted in triplicate. Following incubation, the wells were washed, fixed, stained, and analyzed as described above. Cultures without extract served as growth controls, and TSB alone served as a blank.
The percentage of biofilm inhibition was calculated as follows:
OD Inhibition (%) = (1 − ODGC / ODx) × 100
where ODGC is the mean optical density of the growth control, and ODx is the mean optical density at the tested concentration as previously described.19
Statistical analysis
Biofilm-inhibitory activity (%) was compared between agrII-positive and agrII-negative MRSA isolates at each concentration using Welch’s two-sample t-test (GraphPad Prism v10.2.3), with significance set at P < 0.05. Normality within groups was evaluated using the Shapiro-Wilk test, and homogeneity of variance was assessed using the F-test. Nonlinear regression using the four-parameter log-logistic (LL.4) model (DRC package, R v4.4.1) was applied to generate dose-response curves characterizing genotype-specific biofilm inhibition profiles.
Antimicrobial resistance profile of clinically isolated MRSA
The antimicrobial susceptibility profiles of the 10 Staphylococcus aureus isolates confirmed their methicillin-resistant phenotype (Figure 1). All isolates were resistant to cefoxitin (FOX), penicillin (PEN), and oxacillin (OXA), and most were resistant to trimethoprim-sulfamethoxazole (SXT). Variable resistance was observed for erythromycin (ERY) and clindamycin (CLI), whereas vancomycin (VAN), linezolid (LZD), tetracycline (TET), rifampicin (RIF), and ciprofloxacin (CIP) remained effective. These resistance patterns are consistent with the phenotypic characteristics of methicillin-resistant Staphylococcus aureus (MRSA) and were used as an initial screening step, with molecular confirmation (mecA detection) presented in subsequent analyses.
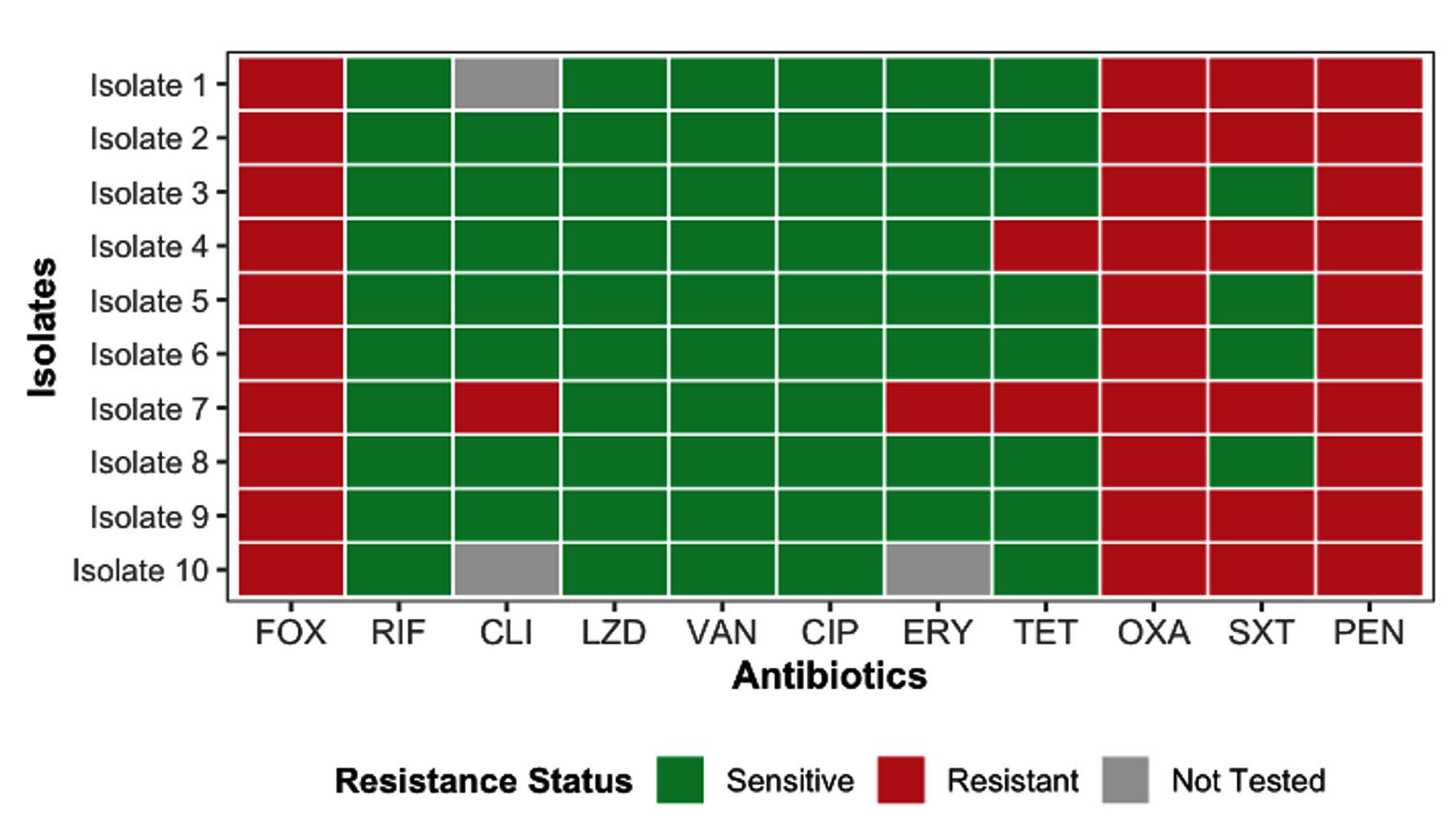
Figure 1. Antimicrobial Resistance Profile of the Clinically Isolated MRSA
Although antimicrobial resistance in S. aureus has been associated with biofilm formation in previous studies, no direct association was evaluated in the present study, and the data are presented as descriptive findings. These findings served as the basis for assessing the biofilm-forming capability and genotypic profiles of the isolates in subsequent analyses.
Biofilm-associated gene profiles of MRSA isolates
The phenotypic biofilm formation capability of the ten MRSA isolates was predominantly moderate (0.140 < OD < 0.280), with two isolates (3 and 10) classified as strong (OD > 0.280). However, marked differences were observed in the biofilm-associated gene profiles. Representative agarose gel electrophoresis confirming PCR amplification of selected biofilm-associated genes (sarA) is shown in Figure 2. The expected amplicon size (~275 bp) was observed in the positive control, whereas no amplification was detected in the negative control, confirming assay specificity. All isolates possessed mecA, confirming phenotypic methicillin resistance. The biofilm-associated genes icaA, icaD, and sarA were detected in all isolates, whereas bap was not detected. Despite the absence of bap, moderate to strong biofilm formation was observed, consistent with ica-dependent polysaccharide-mediated biofilm formation. Notably, five isolates carried agrII, whereas the remaining five were negative (Figure 3a), indicating variability in the quorum-sensing profiles that may influence biofilm regulation rather than initial formation.
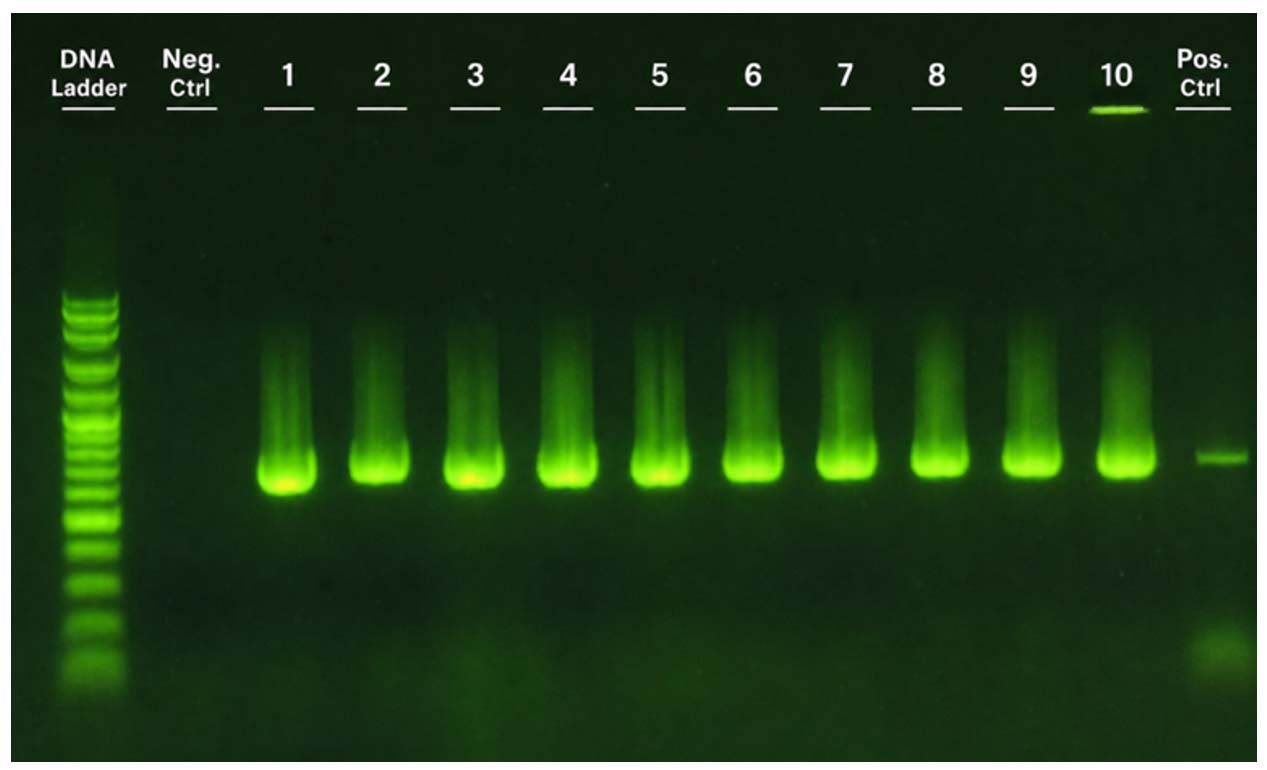
Figure 2. Agarose gel electrophoresis (1.5%) showing PCR amplification of the sarA gene across samples 1-10, including DNA ladder, negative control, and Staphylococcus aureus ATCC 25923 as the positive control, visualized using the GELATOTM system (miniPCR Bio, Cambridge, MA, USA)
Antibiofilm activity of P. amboinicus extract
To determine whether this genotypic variation influenced antibiofilm susceptibility, the activity of the P. amboinicus ethanolic extract was assessed against these isolates (Figure 3b). Isolates carrying agrII exhibited lower susceptibility to biofilm inhibition (12.92% ± 7.45%) compared to agrII-negative isolates, which showed higher inhibition (32.98% ± 1.18%) at 100% concentration. These trends are supported by the numerical values presented in Table 2. The observed antibiofilm activity is likely attributable to the phenolic, flavonoid, and tannin contents of the extract. Moreover, the differential response suggests that agrII may modulate biofilm responsiveness to the extract, consistent with its role in quorum sensing and matrix formation stability.
Table (2): Biofilm Inhibition by Plectranthus amboinicus Extract across agrII genotypes
Concentration (% vol/vol) |
AgrII positive genotypes (Mean ± SD), n = 5 |
AgrII negative genotypes (Mean ± SD), n = 5 |
t-statistic |
P-value |
|---|---|---|---|---|
100 |
12.9 ± 7.4 |
33.0 ± 1.2 |
-5.947 |
0.0034 |
50 |
13.7 ± 12.5 |
23.0 ± 7.2 |
-1.435 |
0.1984 |
25 |
2.8 ± 4.1 |
10.4 ± 6.8 |
-2.122 |
0.074 |
12.5 |
8.2 ± 9.7 |
4.1 ± 4.3 |
0.862 |
0.4244 |
6.25 |
0.6 ± 1.3 |
4.6 ± 5.8 |
-1.516 |
0.1974 |
3.125 |
2.0 ± 4.5 |
4.0 ± 7.9 |
-0.502 |
0.6325 |
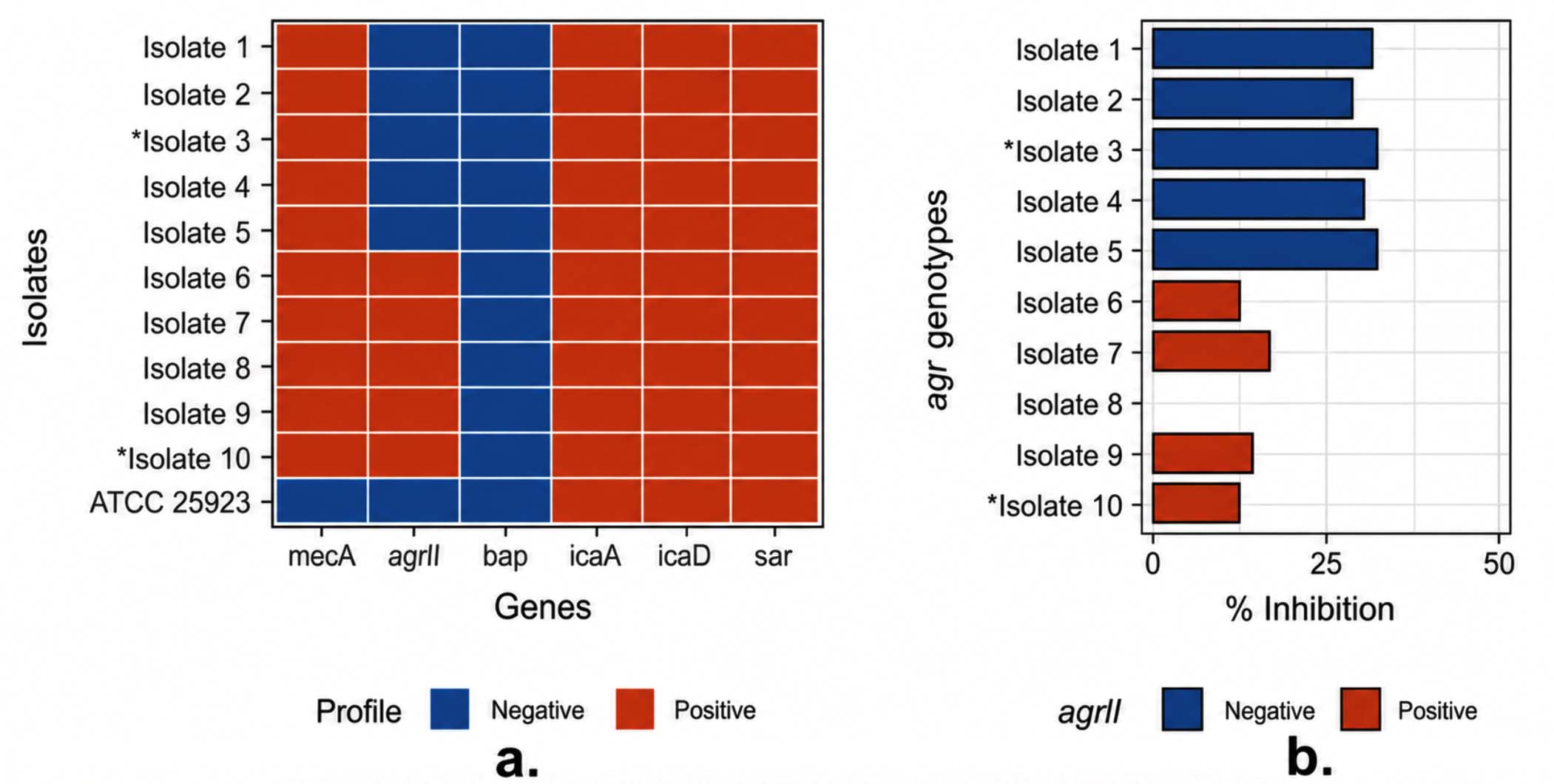
Figure 3. Biofilm-associated gene profile (a) and biofilm-inhibitory activity of P. amboinicus ethanolic extract against clinically isolated MRSA (b). Asterisks (*) mark phenotypically strong biofilm-forming isolates
Dose-dependent antibiofilm activity of P. amboinicus extract
P. amboinicus ethanolic extract demonstrated dose-dependent biofilm inhibition across serial dilutions (100%-3.125% v/v), with agrII-positive strains consistently exhibiting lower levels of inhibition than agrII-negative strains (Figure 4). Nonlinear regression using the four-parameter log-logistic (LL.4) model further illustrated distinct dose-response curves for each genotypic group, with associated 95% confidence bands (shaded blue = agrII-negative; red = agrII-positive) showing minimal overlap across concentrations, where model predictions were reliable (12.5%-100%).
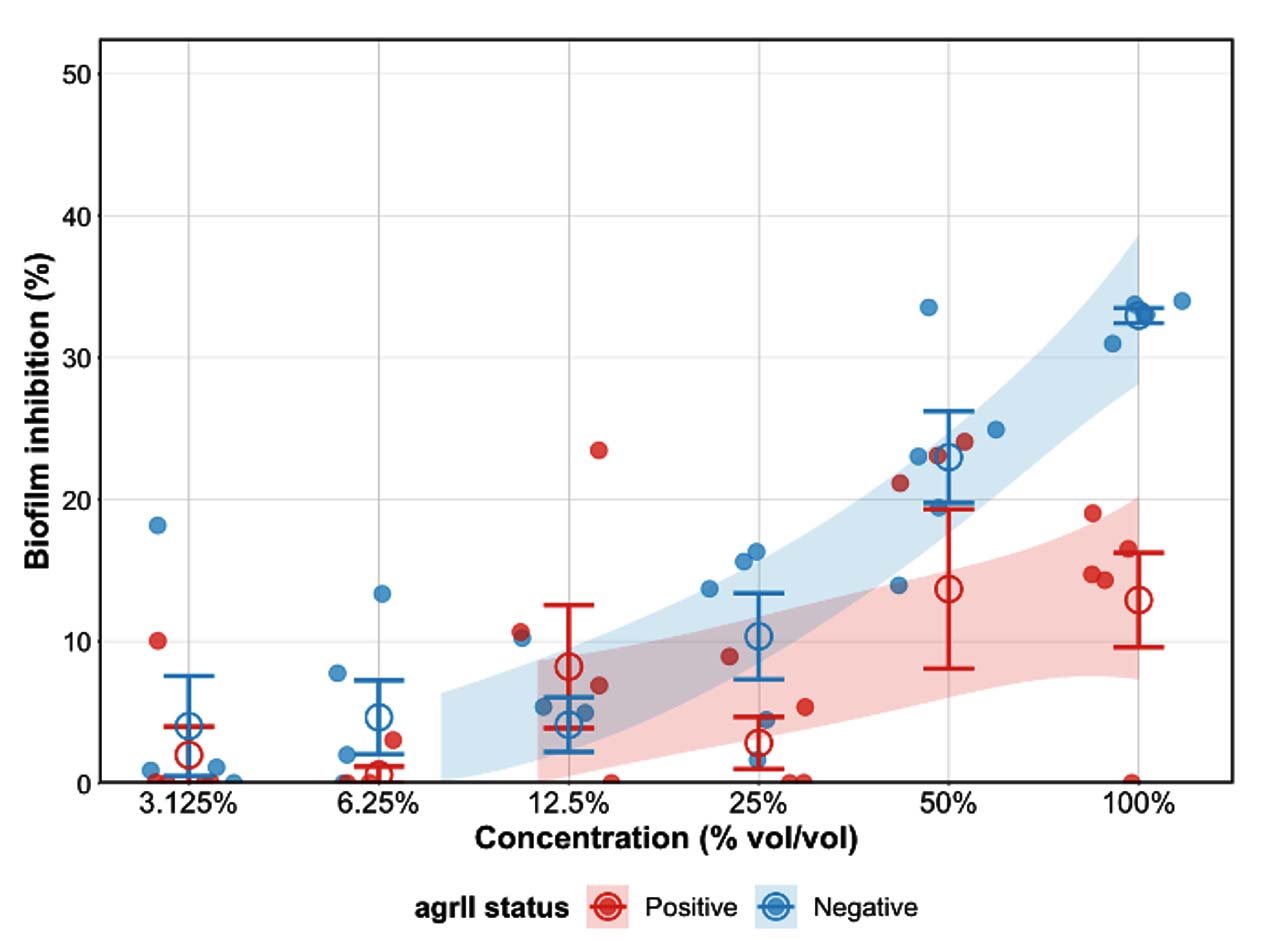
Figure 4. Dose-dependent Biofilm Inhibition of Oregano Ethanolic Extract Against agrII-positive and agrII-negative Biofilm-forming MRSA Genotypes
These trends are supported by the numerical values presented in Table 2. Statistical analysis confirmed that agrII status significantly affected inhibition at 100% concentration
(P = 0.003), whereas the differences were non-significant at lower concentrations (50%: 13.7% ± 12.5% vs. 23.0% ± 7.2%, P = 0.198; 25%: 2.8% ± 4.1% vs. 10.4% ± 6.8%, P = 0.074; 12.5%: 8.%2 ± 9.7% vs. 4.1% ± 4.3%, P = 0.424). However, relatively high standard deviation values at certain concentrations (e.g., 50%) indicate substantial variability among isolates, and these results should be interpreted cautiously.
Nevertheless, given the observed variability and limited sample size, these differences should be considered indicative of trends rather than definitive genotype-specific effects.
Emerging evidence from recent studies have highlighted the central role of biofilm-associated genes and quorum-sensing systems in the persistence and treatment failure of MRSA infections.12,13,18 In particular, the accessory gene regulator (agr) system is a key modulator of biofilm architecture, virulence factor expression, and antimicrobial tolerance.20,21 In this context, the present study contributes to the growing body of work on biofilm-targeted strategies by demonstrating that the ethanolic extract of P. amboinicus exhibits genotype-associated antibiofilm activity in vitro, with agrII-positive MRSA isolates showing reduced susceptibility compared to agrII-negative genotypes at the same concentration of extract.
All clinical isolates carried mecA, confirming their methicillin resistance. The biofilm-associated genes icaA and icaD, which are core components of the intercellular adhesion (ica) operon responsible for polysaccharide intercellular adhesin (PIA) biosynthesis, were detected in all isolates.22 In addition, sarA was present, whereas bap was not detected. These findings indicate that the classical ica-dependent biofilm pathways remain intact in these isolates. While the presence of agrII may be associated with differences in biofilm behavior, its role in modulating biofilm susceptibility cannot be definitively established without gene expression analyses.
The phytochemical profile of P. amboinicus ethanolic extract, characterized by the presence of phenolic compounds, flavonoids, and tannins, provides a plausible basis for the observed antibiofilm activity of the extract. Phenolics, such as carvacrol and thymol, have been reported to interfere with quorum-sensing pathways and disrupt biofilm structure.23 However, quorum-sensing inhibition and gene expression changes were not directly assessed in this study; therefore, mechanistic interpretations should be considered hypothetical.
Comparable antibiofilm effects of phenolic-rich plant extracts have been reported in MRSA and other bacterial systems, supporting the broader potential of natural products as biofilm-disrupting agents in various bacterial systems. The observed genotype-associated differences in inhibition are consistent with previous finding, suggesting that bacterial regulatory systems may influence the responsiveness to antimicrobial compounds.24
The application of a four-parameter log-logistic model to dose-response data provides a quantitative framework for comparing inhibition profiles. However, given the limited sample size and number of data points, these results should be interpreted as indicative of trends rather than definitive genotype-specific effects.
The findings of this study should be interpreted in light of its limitations. The relatively small sample size and in vitro design limit the statistical robustness and generalizability of the findings, and the absence of gene expression analysis and quantitative chemical characterization restricts mechanistic interpretation. In addition, detailed antimicrobial efficacy parameters and safety-related assessments were not performed. These constraints highlight the preliminary nature of the findings and the need for further research.
From a clinical perspective, the potential application of P. amboinicus extract as an antibiofilm agent has yet to be established. Future studies should address the optimal dosing, delivery strategies, safety, and in vivo efficacy of these compounds to determine their therapeutic feasibility.
Overall, while this study provides preliminary evidence of genotype-associated differences in antibiofilm activity, further investigations incorporating molecular, chemical, and translational approaches are required to confirm these findings and elucidate their underlying mechanisms.
This in vitro study demonstrated that the ethanolic extract of P. amboinicus exhibited dose-dependent antibiofilm activity against MRSA isolates, with differences observed between the agrII-positive and agrII-negative genotypes. These findings suggest a potential association between agrII status and variability in antibiofilm response, although this relationship requires further validation. However, these results should be interpreted in light of several limitations, including the small sample size, absence of gene expression analysis, and lack of quantitative chemical and efficacy assessments.
Overall, this study provides preliminary evidence supporting the potential of P. amboinicus extract as an antibiofilm agent, warranting further investigation through mechanistic studies and in vivo validation to assess its clinical applicability.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
Both authors listed have made substantial, direct, and intellectual contributions to the work and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of the University of the Immaculate Conception (protocol code GS-EX-07-25-0058).
- Thacharodi A, Hassan S, Ahmed T, et al. Methicillin-resistant Staphylococcus aureus is raising global concern as it overcomes immune challenges through various virulence mechanisms. iScience. 2026;29(1):114376.
Crossref - Kaushik A, Kest H, Sood M, Steussy B, Thieman C, Gupta S. Biofilm producing methicillin-resistant Staphylococcus aureus (MRSA) infections in humans: Clinical implications and management. Pathogens. 2024;13(1):76.
Crossref - Kot B, Sytykiewicz H, Sprawka I, Witeska M. Effect of trans-Cinnamaldehyde on methicillin-resistant Staphylococcus aureus biofilm formation: Metabolic activity assessment and analysis of biofilm-associated gene expression. Int J Mol Sci. 2019;21(1):102.
Crossref - Kot B, Sytykiewicz H, Sprawka I, Witeska M. Effect of manuka honey on biofilm-associated gene expression during methicillin-resistant Staphylococcus aureus biofilm formation. Sci Rep. 2020;10(1):13552.
Crossref - Silva JM dos S, Almeida JRG da S, Alves C dos SC, et al. Antimicrobial activity from Plectranthus amboinicus (Lour.) Spreng: A review. Eur J Med Plants. 2020;31:1-14.
Crossref - Idrees M, Sawant S, Karodia N, Rahman A. Staphylococcus aureus biofilm: Morphology, genetics, pathogenesis and treatment strategies. Int J Environ Res Public Health. 2021;18(14):7602.
Crossref - Aboelnaga N, Elsayed SW, Abdelsalam NA, et al. Deciphering the dynamics of methicillin-resistant Staphylococcus aureus biofilm formation: From molecular signaling to nanotherapeutic advances. Cell Commun Signal. 2024;22(1):188.
Crossref - Manlubatan SIT, Matias KMH, Mendoza KRP, et al. Determination of the antimicrobial property of oregano (Plectranthus amboinicus (Lour.) Spreng.) crude aqueous leaf extract against throat pathogens using broth and checkerboard dilution methods. Acta Med Philipp. 2022;56:4.
Crossref - Mora-Zuniga AE, Trevino-Garza MZ, Guerra CAA, et al. Comparison of chemical composition, physicochemical parameters, and antioxidant and antibacterial activity of the essential oil of cultivated and wild Mexican oregano Poliomintha longiflora Gray. Plants. 2022;11(14):1785.
Crossref - Guo N, Bai X, Shen Y, Zhang T. Target-based screening for natural products against Staphylococcus aureus biofilms. Crit Rev Food Sci Nutr. 2023;63(14):2216-2230.
Crossref - Coccimiglio J, Alipour M, Jiang ZH, Gottardo C, Suntres Z. Antioxidant, antibacterial, and cytotoxic activities of the ethanolic Origanum vulgare extract and its major constituents. Oxid Med Cell Longev. 2016;2016:1404505.
Crossref - Merritt JH, Kadouri DE, O’Toole GA. Growing and analyzing static biofilms. Curr Protoc Microbiol. 2006.
Crossref - Plota M, Sazakli E, Giormezis N, et al. In vitro anti-biofilm activity of bacteriophage K (ATCC 19685-B1) and daptomycin against staphylococci. Microorganisms. 2021;9(9):1853.
Crossref - Khaleghi M, Khorrami S. Down-regulation of biofilm-associated genes in mecA-positive methicillin-resistant S. aureus treated with M. communis extract and its antibacterial activity. AMB Express. 2021;11(1):85.
Crossref - Novak I, Sipos L, Kokai Z, Szabo K, Pluhar Z, Sarosi S. Effect of the drying method on the composition of Origanum vulgare L. subsp. hirtum essential oil analysed by GC-MS and sensory profile method. Acta Aliment. 2011;40(Suppl 1):130-138.
Crossref - Kumar P, Singh S, Kumar N. Plectranthus amboinicus: A review on its pharmacological and pharmacognostical studies. Am J Physiol Biochem Pharmacol. 2020;10(2):55-62.
Crossref - Pérez-Delgado O, Alvarado-Pineda RL, Yacarini-Martínez AE. In vitro antibacterial activity of crude ethanolic extract from the leaves of Origanum vulgare against Staphylococcus aureus ATCC 29213, Pseudomonas aeruginosa ATCC 27853 and Escherichia coli ATCC 25922. J Selva Andina Res Soc. 2021;12(1):21-29.
Crossref - Amri IA, Ramadani NF, Hamidah F, Dameanti FNAEP, Adrenalin SL. Potential antibacterial effects of ethanol extract and essential oil of Origanum vulgare on Klebsiella pneumoniae and Staphylococcus aureus. World Vet J. 2023;13(4):486-491.
Crossref - Mezzasalma N, Spadini C, Spaggiari C, et al. Antibacterial and antibiofilm activity of Eucalyptus globulus leaf extract, asiatic acid and ursolic acid against bacteria isolated from bovine mastitis. Front Vet Sci. 2025;12:1565787.
Crossref - Tan L, Li SR, Jiang B, Hu XM, Li S. Therapeutic targeting of the Staphylococcus aureus accessory gene regulator (agr) system. Front Microbiol. 2018;9:55.
Crossref - Goldie SP, Lau LC, Jones HAS, Harries PG, Walls AF, Salib RJ. Identification of novel Staphylococcus aureus core and accessory virulence patterns in chronic rhinosinusitis. Int J Mol Sci. 2025;26(8):3711.
Crossref - Nguyen HTT, Nguyen TH, Otto M. The staphylococcal exopolysaccharide PIA: Biosynthesis and role in biofilm formation, colonization, and infection. Comput Struct Biotechnol J. 2020;18:3324-3334.
Crossref - Touati A, Mairi A, Ibrahim NA, Idres T. Essential oils for biofilm control: Mechanisms, synergies, and translational challenges in the era of antimicrobial resistance. Antibiotics. 2025;14(5):503.
Crossref - Hiltunen AK, Savijoki K, Nyman TA, et al. Structural and functional dynamics of Staphylococcus aureus biofilms and biofilm matrix proteins on different clinical materials. Microorganisms. 2019;7(12):584.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.