


ISSN: 0973-7510
E-ISSN: 2581-690X
Herbal remedies and medicinal plants have long been valuable natural resources for pharmaceutical, cosmetic, and perfumery industries. Endophytic fungi, which live symbiotically within plants, offer potential therapeutic benefits. The objective of this study was to isolate and characterize endophytic fungal species from the leaves of Mammea suriga Buch.-Ham. ex Roxb., an evergreen tree endemic to the Western Ghats of India. Leaves were collected, surface-sterilized, and cultured on various agar media to isolate endophytic fungi. The cultured fungal strains were subjected for morphological and molecular identification, which resulted in two fungal isolates: Phyllosticta capitalensis (BGMSB-01) and Diaporthe pseudophoenicicola (BGMSW-01). The extracts of both fungal isolates showed significant inhibition against both Gram-positive Staphylococcus aureus and Gram-negative Escherichia coli. The zones of inhibition indicated that D. pseudophoenicicola (BGMSW-01) has stronger inhibition (3.79 ± 1.48 mm and 2.39 ± 0.67 mm, respectively) compared to P. capitalensis (BGMSB-01) (inhibition zones of 1.2 ± 0.52 mm and 1.72 ± 0.18 mm, respectively). Plant Growth-Promoting (PGP) activity was evaluated using Phaseolus lunatus seeds, where isolate BGMSB-01 completely inhibited the germination, while BGMSW-01 showed significant increase in germination and seedling growth promotion.
Antimicrobial Activity, Diaporthe pseudophoenicicola, Phyllosticta capitalensis, Identification, PGP Activity
Herbal remedies have been used for thousands of years to treat a wide range of diseases and plants remain a key source of natural bioactive compounds. Currently, the pharmaceutical, cosmetic, and perfumery industries are increasingly relying on using medicinal plants and their highly-valuable secondary metabolites as raw materials.1 Like other organisms, plants also host diverse groups of microbes, including endophytes— Endophytic microorganisms inhabit plant tissues internally without inducing disease symptoms or damage. Often these endophytes are emerging as promising sources of novel therapeutic agents which produces or secrete valuable secondary metabolites. In this perspective, medicinal plants are now recognized not only for their pharmacological properties but also as reservoirs of diverse endophytic microorganisms with valuable secondary metabolites with significant biotechnological potential.2,3
In the current study, leaves of Mammea suriga (Buch.-Ham. ex Roxb.), a medicinal tree from Karnataka, India, was screened for the endophytic fungi. This species is endemic to the Central Western Ghats of India. It holds economic importance in the dye and perfume industries and is traditionally used in treating various ailments.4 Local healers use its flower buds to treat skin eruptions, itching, sore throats, coughs, vomiting, and diarrhea, while stem bark extracts are employed to manage fever, foul breath, bleeding piles, and other conditions.4,5 In an effort to identify the endophytic fungal flora from the medicinal plants of the region, this research focused on the isolation and identification of endophytic fungi from leaves of M. suriga, evaluating their antimicrobial and plant growth-promoting activities.
Collection of plant material
Leaves of M. suriga were collected from the ICMR-National Institute of Traditional Medicine campus in Belagavi, India (15°512 N, 74°302 E; elevation: 752 m above sea level). A plant taxonomist authenticated the collected specimen and has been deposited in the herbarium of ICMR-NITM, Belagavi (Voucher Number: RMRC 1716). The region experiences a tropical climate with an average annual rainfall of 1468 mm (57.8 inches) (https://en.climate-data.org/asia/india/karnataka/belagavi-4076). The collected leaves were immediately transported to the laboratory for further processing.
Isolation of endophytic fungi
Healthy M. suriga leaves underwent initial cleaning by washing under running tap water for 30 minutes, followed by rinsing with distilled water for 15 minutes.6 Surface decontamination involved immersion in 1% Tween 20 solution for 2 minutes to remove epiphytic microbes, with subsequent thorough rinsing using double-distilled water.7 Endophytic fungi were isolated from these leaves using established protocols with minor modifications.8 The leaf samples were immersed in 70% ethanol (2-3 min), then treated with 4% sodium hypochlorite solution (2-5 min) for surface sterilization, followed by thorough rinsing with distilled water and aseptic air-drying.9 The leaf imprint method was used to assess the effectiveness of surface sterilization.10 Using a sterile blade, the samples were cut into 0.5 × 0.5 cm squares. Plant tissue segments (two per plate) were placed on potato dextrose agar (PDA), nutrient agar (NA), and Sabouraud dextrose agar (SDA) amended with 150 mg/L streptomycin sulfate to recover endophytic fungi.9 All procedures were performed under sterile conditions. The plates were incubated at 26 ± 2 °C. Actively growing fungal colonies emerging from the leaf tissues were subculture onto PDA, NA, and SDA plates for further identification.11
Extract preparation
Healthy leaf samples were collected, shade-dried and ground into coarse powder. Two grams of leaf powder was treated with 30 mL of 0.01 M sulfuric acid (H2SO4) and incubated at 25 °C with continuous shaking at 170 RPM for 30 min. Subsequently, 7.5 mL of 0.4 M sodium bicarbonate (NaHCO3) was added to the mixture, and shaking was continued at the same speed for an additional 60 min. The extract was then filtered using Whatman No. 4 filter paper and concentrated in a water bath at 55 °C. The dried extract was weighed and stored in airtight containers at 4 °C until further use.12
Culture and Preparation of Fungal Biomass
Endophytic fungal isolates were cultured individually in 500 mL of Sabouraud Dextrose (SD) broth prepared in 1000 mL conical flasks. Each flask was inoculated separately with a pure culture of the respective endophytic fungus under sterile conditions. The cultures were incubated for 30 days at ±27 °C under static conditions. After the incubation period, the mycelial biomass was harvested by filtration and dried. The dried mycelia were ground into a fine powder using a sterile mortar and pestle, and the powdered biomass was stored in airtight containers under dry conditions for further analysis.13
Extraction of fungal mycelia
From the stored mycelial powder, 0.25 g was mixed with 3.75 mL of 0.01M H2SO4 in a flask and was kept for continuous shaking on an orbital shaker (Orbitek, Scigenics Biotech India) for 30 min. Then 0.93 mL of 0.4 m NaHCO3 was added to the flask and the shaking was continued at a speed of 170 rpm in a controlled temperature (25 °C) for 60 min. The resultant extract was filtered through Whatman’s filter paper No. 4, and the filtrate was kept for drying on water bath at 55 °C. The dried powder was stored for further use.9
DNA extraction and sequencing
DNA extraction was performed using the standard protocols provided with the DNA isolation kits (Sigma-Aldrich, India), with minor modifications. Genomic DNA was extracted from both leaf samples and fungal isolates.
The extracted DNA was sequenced using the Illumina NovaSeq 6000 platform, generating approximately 5 GB of raw sequencing data. The raw reads were processed using FastP14 for quality filtering and adapter trimming. Sequence quality was assessed using FastQC, and the results were summarized using MultiQC.15 De novo genome assembly was performed using the AAFTF pipeline, a fungal genome assembler.16 Assembly refinement and error correction were carried out using the built-in tools available within the same pipeline. The internal transcribed spacer (ITS) subregion was subsequently extracted from the assembled genome using ITS × v1.1.3.17
Morphological and Molecular identification
Morphological characterization of the fungal isolates was performed based on microscopic examination and colony characteristics, including colony color, filamentous growth, and mat type. The fungal strains were stained with Lactophenol Cotton Blue and observed under a Magnus MX-21i LED FS11 microscope at 40× and 100× magnifications. Structural features such as mycelial arrangement, fruiting bodies, conidia, conidiophores, and reproductive and non-reproductive structures were examined.18 Standard fungal identification keys were used for morphological classification.19
Phylogenetic analysis
The ITS sequences were used for phylogenetic analysis. Reference ITS sequences of closely related fungal species within the same genus and family were retrieved from the NCBI database, along with Agaricus bisporus (phylum Basidiomycota) as an outgroup. Multiple sequence alignment was performed using ClustalW integrated in MEGA XI.20 A phylogenetic tree was constructed using the neighbor-joining (NJ) method with 1,000 bootstrap replicates to assess branch support.
Antibacterial assay
The antibacterial activity of the extracts was assessed using the agar well diffusion method.21-23 The sterile (Mueller-Hinton Agar, Himedia) plates were prepared appropriate for each test organism and were inoculated with the test organisms Staphylococcus aureus and Escherichia coli. Six mm diameter wells were made in the media using a sterile cork borer. 20 µL of the extract was added to each well. Streptomycin (Hi-Media) was used as the positive control for bacteria. Similarly, 20 µL methanol was tested as negative control. The plates were then incubated at 35 °C for 24 hrs. The diameter of the clear inhibition zone around the well was measured using a ruler. All antibacterial assays were performed in triplicate, and results were expressed as mean ± SD. Statistical analysis was carried out using unpaired two-tailed Student’s t-test with
P < 0.05 considered statistically significant.
PGP activity
The experiment was conducted using Lima bean (Phaseolus lunatus L.) seeds. The seeds selection was made by sinker and floater technique.24 The seeds were surface sterilized by first washing with running water followed by treatment with 40% tween 20 solution. It was followed by treatment with 4% sodium hypochlorite solution for 3 min, followed by rinsing with sterile water.25 Six seeds of uniform size and weight were soaked in the mycelial grown broth for 8 hours, while those soaked in distilled water and sterile broth were the control treatment. The seeds of each treatment were placed on a moist filter paper in sterile Petri dishes. The filter papers in the Petri dishes were kept moist distilled water throughout the experiment to maintain adequate moisture for seed germination. Petri dishes were incubated at 25 °C for 9 days. The seeds were observed daily from day 3 to day 9, for their germination and growth. The plant growth-promoting assay was conducted in triplicate, and growth data were expressed as mean ± SD.
Isolation and identification of endophytic fungi
Two endophytic fungi were isolated from the leaves of M. suriga. The cultures were evaluated for colony morphology, and microscopic examination of spores and mycelia (Figure 1). Based on the morphology and microscopy, the first culture (BGMSB-01) was identified as Phyllosticta capitalensis and the second culture (BGMSW-01) as Diaporthe pseudophoenicicola.
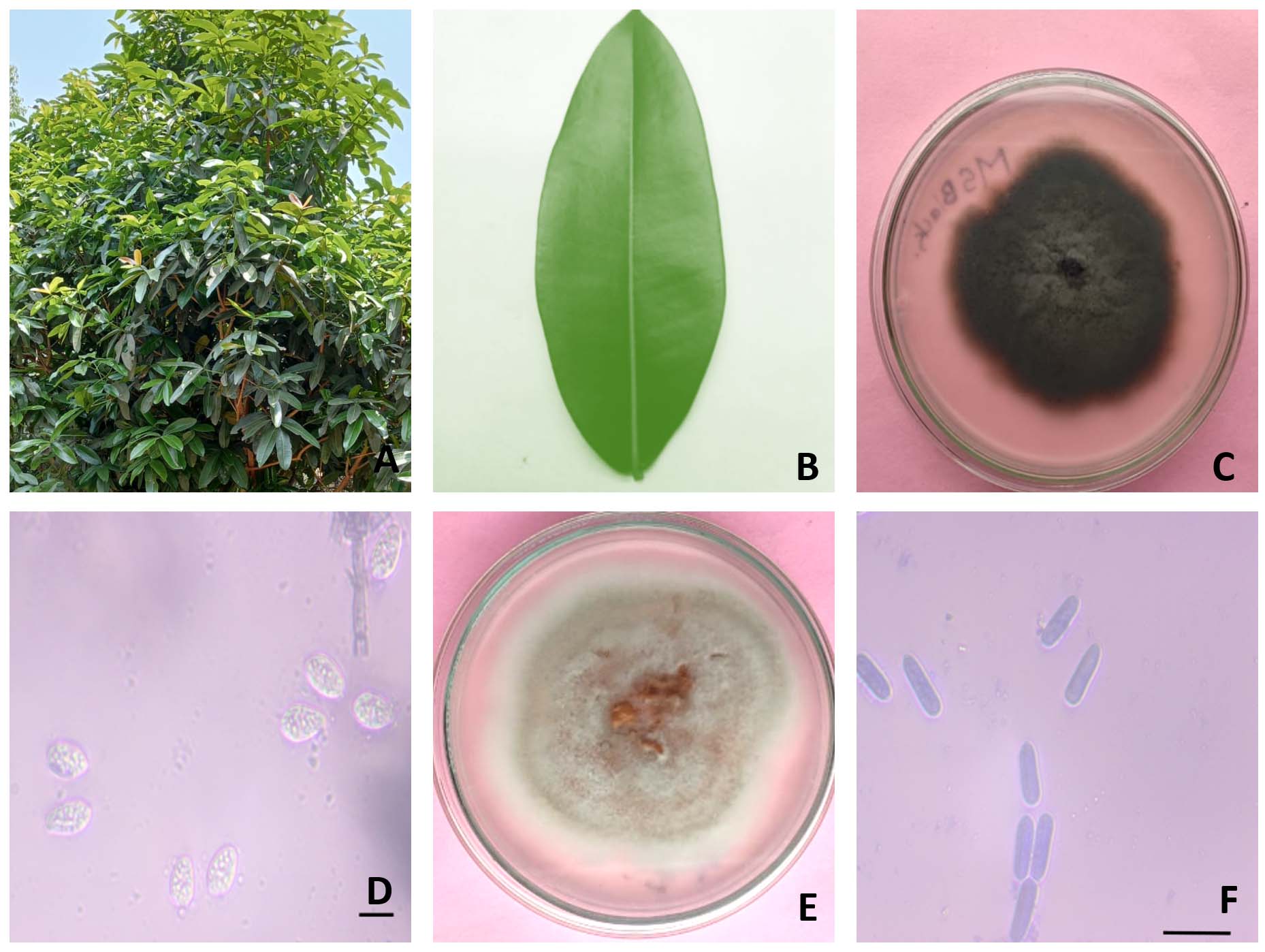
Figure 1. Morphological characteristics of endophytic fungi isolated from Mammea suriga leaves. (A) Habit of M. suriga plant; (B) Leaf sample used for fungal isolation; (C) Colony morphology of Phyllosticta capitalensis (BGMSB-01) on culture medium; (D) Microscopic view of conidia of P. capitalensis; (E) Colony morphology of Diaporthe pseudophoenicicola (BGMSW-01); (F) Microscopic view of conidia of D. pseudophoenicicola
In the molecular identification, the sequenced ITS region contained ITS1, 5.8S rRNA and ITS2 regions. Further, these sequences were used for nucleotide BLAST in NCBI database. The ITS sequence of BGMSB-01 isolate showed 100% sequence similarity to Phyllosticta capitalensis, the closest species and BGMSW-01 sequence showed 99.82% similarity to the closest species, Diaporthe pseudophoenicicola. The ITS sequences were uploaded to NCBI database with accession numbers PQ493703 for BGMSB-01 and PQ493729 for BGMSW-01 (Table 1).
Table (1):
The ITS sequence identification of the endophytic fungal strains
No. |
Fungal Strain Code |
GenBank |
Accession Number |
Homolog Sequences |
Sequence Identity % |
Closest species Accession Number |
|---|---|---|---|---|---|---|
01. |
BGMSB-01 |
SUB14806299 |
PQ493703 |
Phyllosticta capitalensis strain LCM 826.01 |
100% |
MF495391.1 |
02. |
BGMSW-01 |
SUB14806458 |
PQ493729 |
Diaporthe GM143 Pseudophoenicicola isolate |
99.82% |
OR416452.1 |
Phylogenetic analysis
To confirm the identification of fungal strains and their taxonomic and phylogenetic placement relative to other species, a neighbor-joining tree was generated using 1,000 bootstrap replicates along with previously characterized ITS sequences of closely associated species. The tree separated the sequences into different clades and accurately placed along with the ITS sequences of identified species Phyllosticta capitalensis for and BGMSB-01 and Diaporthe pseudophoenicicola for BGMSW-01 (Figure 2).
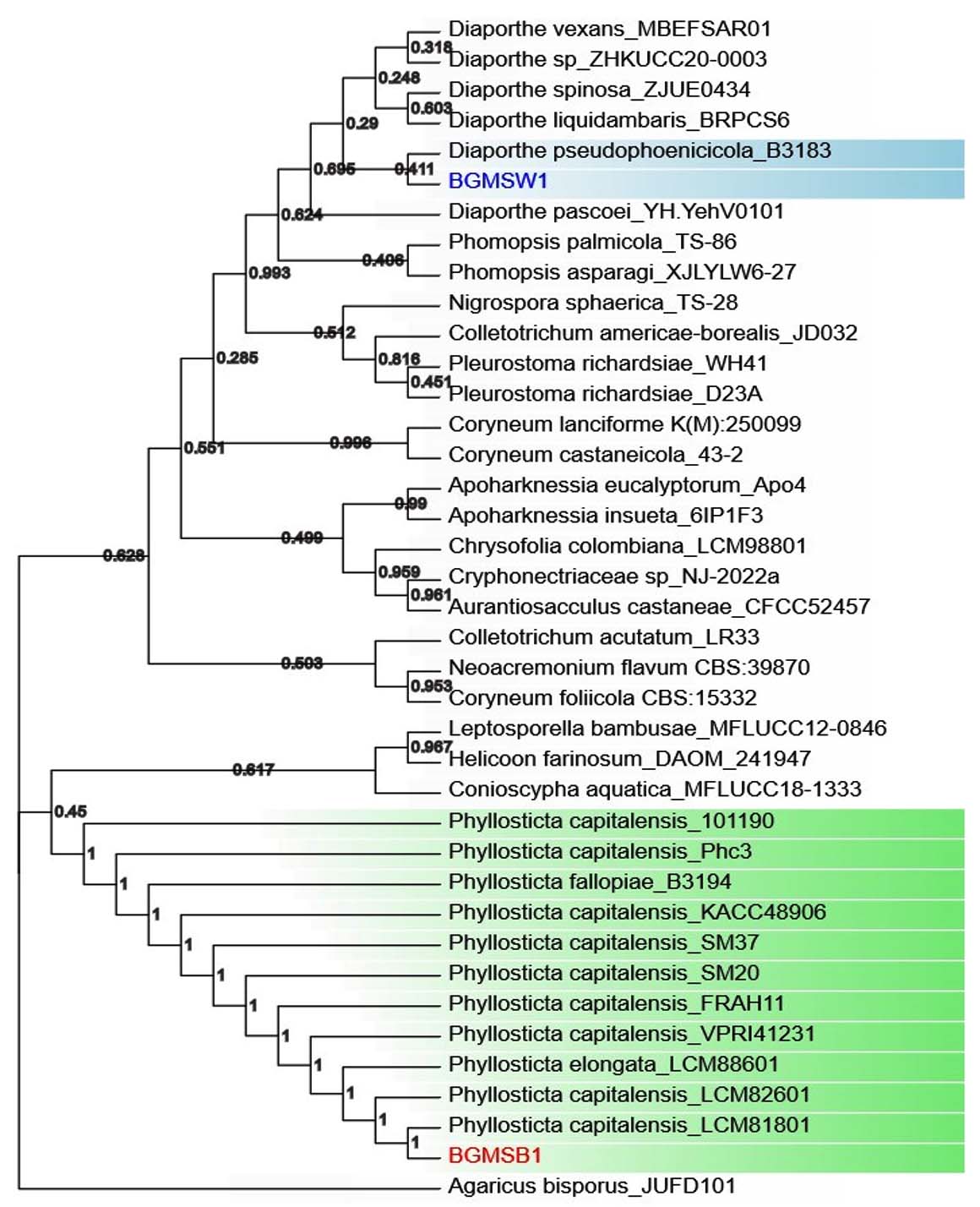
Figure 2. Phylogenetic tree (Neighbor-joining Method with 1000 bootstraps) using ITS gene sequences of fungal isolated BGMSW-01, BGMSB-01 along with ITS sequences of closely related fungal strains
Antimicrobial activity of fungal endophytes
The antimicrobial activity of the endophytic fungal isolates Phyllosticta capitalensis (BGMSB-01) and Diaporthe pseudophoenicicola (BGMSW-01) was evaluated against Staphylococcus aureus and Escherichia coli using the agar well diffusion method. For the Gram-positive bacterium Staphylococcus aureus, the BGMSB-01 extract exhibited weaker antibacterial activity, producing a mean inhibition zone of 1.20 ± 0.52 mm. In contrast, the BGMSW-01 extract demonstrated a relatively larger inhibition zone of 3.79 ± 1.48 mm, suggesting higher antibacterial potential against the Gram-positive strain.
In the case of the Gram-negative bacterium Escherichia coli, the BGMSB-01 extract showed an inhibition zone of 1.72 ± 0.18 mm, while the BGMSW-01 extract again produced a greater zone of inhibition (2.39 ± 0.67 mm) (Table 2). Overall, these results indicate that the extract of isolate BGMSW-01 exhibited comparatively higher inhibitory activity than BGMSB-01 against both bacterial strains, although the inhibition zones were relatively small. It was also observed that BGMSW-01 showed greater variability in inhibition zones across replicates.
Table (2):
Antimicrobial activity of endophytic fungi isolated from Mammea suriga
| No. | Endophytic Fungal Isolates | Zone of inhibition of microorganisms (mm) | |
|---|---|---|---|
| Staphylococcus aureus | Escherichia coli | ||
| 1. | BGMSB-01- Phyllosticta capitalensis | 1.2 ± 0.52 mm | 1.72 ± 0.18 mm |
| 2. | BGMSW-01- Diaporthe pseudophoenicicola | 3.79 ± 1.48 mm | 2.39 ± 0.67 mm |
These findings suggest that both fungal extracts may contain antibacterial metabolites, with BGMSW-01 showing relatively stronger activity, particularly against the tested Gram-positive bacterium.
Evaluation of endophytic fungal strains for their plant growth-enhancing potential
The plant growth-regulating effects of fungal extracts from BGMSB-01 and BGMSW-01 were evaluated over nine days, with water and SDA broth serving as controls. Plant growth, measured as shoot length (in cm), was recorded daily, and the results were tabulated as (mean ± SD) (Table 3). The extract of Phyllosticta capitalensis (BGMSB-01) isolate showed a consistent growth-suppressing effect with no seedling germination observed until day 9 of observation. This indicates a strong inhibitory effect of the extract on plant growth or its ability to potentially induce or maintain dormancy.
Table (3):
Endophytic Fungal Strains as Plant Growth-Promoting
Day |
Water (in cm) |
Control (SDA Broth) |
Phyllosticta capitalensis cultured broth |
Diaporthe pseudophoenicicola cultured broth (in cm) |
|---|---|---|---|---|
Day 3 |
1.3 ± 0.63 |
0.76 ± 0.058 |
0 |
1.32 ± 1.16 |
Day 4 |
1.73 ± 0.71 |
1.1 ± 0.1 |
0 |
1.87 ± 1.56 |
Day 5 |
2.07 ± 0.68 |
1.46 ± 0.25 |
0 |
2.48 ± 1.53 |
Day 6 |
2.42 ± 0.64 |
1.86 ± 0.32 |
0 |
3.3 ± 1.85 |
Day 7 |
2.67 ± 0.58 |
2.23 ± 0.51 |
0 |
3.8 ± 2.11 |
Day 8 |
3.12 ± 0.45 |
2.83 ± 0.5 |
0 |
3.97 ± 2.2 |
Day 9 |
3.53 ± 0.37 |
3.33 ± 0.45 |
0 |
4.37 ± 2.5 |
In contrast, the extract of isolate Diaporthe pseudophoenicicola (BGMSW-01) promoted seedling growth substantially. On Day 3, the treated seedlings showed a growth of 1.32 ± 1.16 cm, which was comparable to the water control (1.30 ± 0.63 cm) but much higher than the SDA broth control (0.76 ± 0.058 cm). From Day 4 onwards, the growth of plants treated with BGMSW-01 extract consistently outpaced that of both controls. By Day 9, seedlings treated with BGMSW-01 had reached a mean height of 4.37 ± 2.5 cm, compared to 3.53 ± 0.37 cm for the water control and 3.33 ± 0.45 cm for the SDA broth control. Representative photographic records of seed germination and seedling growth under different treatments were documented during the experiment and are provided as Supplementary Figure S1.
These results suggest that the extract of Diaporthe pseudophoenicicola (BGMSW-01) isolate enhances plant growth compared to both water and SDA broth controls, while Phyllosticta capitalensis (BGMSB-01) isolate completely inhibited seedling germination. The study concluded that the extract of Phyllosticta capitalensis (BGMSB-01) isolate is a potent germination and growth inhibitor, while Diaporthe pseudophoenicicola (BGMSW-01) extract promotes enhanced plant growth, making it a potential bio-stimulant for agricultural applications.
Endophytic fungi represent a very important underexplored domain within plant microbiome. They involve and influence the host plant physiology through diverse mechanisms, including the production of bioactive secondary metabolites, regulation of host gene expression, modulation of host defense responses, and alterations in growth patterns.2 They also reported to produce valuable secondary metabolites. They are also reported to produce high value secondary metabolites produced by their host plants. For example, an endophytic fungus isolated from the fruits of Taxus cuspidata, a plant which produce chemotherapeutic drug Taxol, is also capable of producing Taxol under culture.26 Exploring the endophytic fungi from medicinal plants can get novel strains of fungi that may produce high value therapeutically relevant metabolites and is also quickly culturable with optimized conditions and can be manipulated by genetic engineering to produce to increase yield.27
In this study, two endophytic fungi isolated from the leaves of M. suriga, Phyllosticta capitalensis (BGMSB-01) and Diaporthe pseudophoenicicola (BGMSW-01), were systematically characterized for their morphological, molecular, antimicrobial and phytoregulatory, revealing stark functional dichotomies between the two isolates.
Molecular identification and phylogenetic placement
The identification of both fungal isolates was done morphological methods and molecular characterization. Molecular identification was done using ITS sequence region and subsequent similarity analysis using NCBI nucleotide BLAST analysis confirmed their species-level affiliations with P. capitalensis and D. pseudophoenicicola, respectively. The microscopic characteristics of mycelia and spores were consistent with the species identified through molecular analysis. The phylogenetic reconstruction based on ITS sequences robustly supported this classification, showing discrete clustering of the isolates with high bootstrap values alongside their closest reference sequences. Such accurate placement is essential, given the high degree of morphological plasticity observed among endophytic fungal genera, particularly within Phyllosticta and Diaporthe, both of which are known to exhibit substantial inter- and intraspecies variation.28,29 The fungal species Phyllosticta capitalensis and Diaporthe pseudophoenicicola are well established to be endophytic in nature in and have been isolated from several plant species.11
Antimicrobial potential and differential bioactivity
The endophytic fungi have been reported to be involved in protection of plants from various bacterial and fungal diseases possibly by modulating plant defense mechanisms of by producing various antimicrobial compounds.30 The two isolates displayed markedly different antibacterial activities. Diaporthe pseudophoenicicola (BGMSW-01) showed significantly higher inhibitory effects against both S. aureus and E. coli, with a higher mean inhibition zone. In contrast, Phyllosticta capitalensis (BGMSB-01) exhibited only mild to moderate activity. These results align with earlier studies highlighting the genus Diaporthe as a producer of bioactive compounds, including antimicrobial terpenoids, terpenoids, and flavonoids.12,31 On the other hand, while Phyllosticta spp. are more frequently associated with plant pathogenicity or latent endophytism, there are fewer reports of potent antimicrobial metabolite production.32,33
The superior antibacterial performance of Diaporthe pseudophoenicicola (BGMSW-01) to both Gram-positive and Gram-negative bacteria might be attributed to a richer or more potent profile of antimicrobial metabolites, possibly functioning via membrane disruption, inhibition of nucleic acid synthesis, or reactive oxygen species (ROS) induction mechanisms.34 Furthermore, the greater variability in inhibition zones observed with Diaporthe pseudophoenicicola (BGMSW-01) suggests either heterogeneity in metabolite diffusion or synergistic effects among multiple compounds in the extract.
Impact on seed germination and plant growth
A fascinating divergence in plant-interactive behavior was observed in the germination assays using Phaseolus lunatus seeds. The extract from P. capitalensis (BGMSB-01) demonstrated strong growth-suppressive effects, completely inhibiting seed germination over a 9 days period. There are reports of some fungal metabolites that can affect seed germination.35 Also, this inhibitory potential could be attributed to allelochemicals or growth-suppressing agents such as lactones, quinones, or alkaloids that may interfere with hormone signaling or cellular respiration.36-38
Conversely, the extract of D. pseudophoenicicola (BGMSW-01) significantly enhanced seedling growth, particularly from Day 4 onwards surpassing both water and broth controls. This result suggests the presence of bioactive compounds with phytohormone-like effects, or volatile organic compounds that may stimulate cell division and elongation. Endophytes from Diaporthe are known to synthesize IAA and promote host plant growth under various conditions, enhancing both shoot and root development.39,40 Thus, BGMSW-01 emerges as a potential plant growth-promoting endophyte (PGPE) with applications in sustainable agriculture.
The starkly contrasting effects of the two endophytic isolates-ranging from strong growth suppression to pronounced growth promotion—underscore the functional diversity within endophytic fungal communities. These outcomes have substantial implications for ecological dynamics, including seed germination timing, plant health under biotic or abiotic stress, and microbe-microbe competition within the host.41,42 From a translational standpoint, the antimicrobial potential of isolated strain of D. pseudophoenicicola (BGMSW-01) could be harnessed for developing biocontrol agents or natural antimicrobial formulations, while its plant growth-stimulating properties may be of direct relevance to sustainable agriculture and biofertilizer development.
In conclusion, endophytic fungal isolates showed contrasting effects—one with strong growth suppression activity to the other enhanced growth promotion -highlighting diverse effect or roles of endophytes in plants. These differences can have substantial implications for timing of seed germination, plant growth under biotic or abiotic stress, and microbe-microbe competition within the host and in defense.43 In practical terms, the antimicrobial potential of isolated strain of D. pseudophoenicicola (BGMSW-01) could be used to make developing biocontrol agents or natural antimicrobial formulations, while its plant growth-stimulating properties may also enhance the plant growth.
Meanwhile, the germination-suppressing properties of isolated P. capitalensis (BGMSB-01) strain could find use in weed control or germplasm conservation scenarios. Further exploration of its secondary metabolite profile, particularly the identification of dormancy-inducing compounds, may reveal novel biochemical pathways or molecular mechanisms of seed physiology regulation.
Our findings provide strong evidence that endophytic fungi serve dual ecological roles as regulators of plant growth and as producers of bioactive antimicrobial compounds. The distinct biological activities observed in Phyllosticta capitalensis and Diaporthe pseudophoenicicola highlight the rich, untapped potential of endophytes associated with native plant species such as M. suriga. To further elucidate the molecular basis of these interactions, future studies should include whole-genome sequencing and transcriptomic analysis under co-culture and plant-interaction conditions. Such bioprospecting approaches may lead to uncovering new bioactive molecules with promising uses in both agriculture and healthcare.
Additional file: Figure S1.
ACKNOWLEDGMENTS
The authors are grateful to the Director, Indian Council of Medical Research- National Institute of Traditional Medicine, Belagavi, and the Department of Health Research, Ministry of Health and Family Welfare, Govt. of India, for supporting the study and for providing the facilities.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
HVH conceptualized and supervised the study. MLM and HVH applied methodology. PSK performed Investigation and formal analysis. MLM performed validation. SIJ and KP performed data curation. PSK wrote the original draft. SIJ, HVH, KP and MLM wrote, reviwed and revised the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
This work was supported by the Institutional Grant from the Indian Council of Medical Research-National Institute of Traditional Medicine, Belagavi.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript and/or in the supplementary files.
ETHICS STATEMENT
Not applicable.
- Kumar S, Kaushik N. Endophytic fungi as sources of bioactive metabolites. Microbiol Res. 2021;242:126606.
Crossref - Mishra VK, Passari AK, Chandra P, Leo VV, Kumar B, Uthandi S, Thankappan S, Gupta VK, Singh BP. Endophytes as sources of bioactive compounds: A review. Front Microbiol. 2020;11:599.
Crossref - Aly AH, Debbab A, Kjer J, Proksch P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 2010;41(1):1-16.
Crossref - Bhat KG, Hegde GR. Ethnomedicinal plants of the Western Ghats of Karnataka, India. J Econ Taxon Bot. 2014;38:1-10.
- Nadkarni KM, ed. Indian Materia Medica. 3rd ed. Mumbai: Popular Prakashan; 2000.
- Khalil AMA, Hassan SED, Alsharif SM, et al. Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting. Biomolecules. 2021;11(2):140.
Crossref - Zhang Q, Xiang J, Zhang L, Wang Y. Isolation, identification, and antibacterial evaluation of endophytic fungi from Pinellia ternata. Front Microbiol. 2023;14:1172629.
Crossref - Gowthaman MK, Mohanapriya R. Isolation of endophytic fungi with antimicrobial activity from medicinal plants. J Appl Pharm Sci. 2023;13(9):001-010.
Crossref - Gond SK, Verma VC, Mishra A, Kumar A, Kharwar RN. Role of fungal endophytes in plant protection and their isolation and characterization. Microbiol Res. 2012;167(7):493-501.
Crossref - El-Nageribi SAF, Elshafi AE, Alkhanjari SS. Endophytic fungi associated with Ziziphus species from mountainous area of Oman and new records. Biodiversitas. 2013;14(1):10-16.
Crossref - Bashir A, Manzoor MM, Ahmad T, et al. Endophytic fungal community of Rosa damascena Mill. as a promising source of indigenous biostimulants: Elucidating its spatial distribution, chemical diversity, and ecological functions. Microbiol Res. 2023;276:127479.
Crossref - Besharati M, et al. Extraction and identification of bioactive metabolites from endophytic fungi and evaluation of their antimicrobial activity. Molecules. 2025;30:3870.
Crossref - Khadka G, et al. Isolation and characterization of fungal endophytes from medicinal plants and evaluation of their antimicrobial activity. J Fungi. 2023;9:100
- Chen S, Zhou Y, Chen Y, Gu J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34(Suppl 1):i884-i890.
Crossref - Ewels P, Magnusson M, Lundin S, Kaller M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics. 2016;32(19):3047-3048.
Crossref - Stajich JE, Palmer JM. AAFTF: Automated assembly for the fungi. Microbiol Resour Announc. 2019;8(4):e01619-18.
Crossref - Bengtsson-Palme J, Ryberg M, Hartmann M, et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol Evol. 2013;4(10):914-919.
Crossref - Chowdhury S, Lata R, Gond SK. Bioactivity of fungal endophytes associated with ethno-medicinal plants used by tribal communities of Achanakmar-Amarkantak Biosphere Reserve. J Mycopathol Res. 2022;60:99-110.
- Watanabe T, ed. Pictorial atlas of soil and seed fungi: Morphologies of cultured fungi and key to species. 3rd ed. Boca Raton: CRC Press; 2010.
- Tamura K, Stecher G, Kumar S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38(7):3022-3027.
Crossref - Asfa B, et al. Evaluation of antimicrobial activity of selected medicinal plant extracts using agar well diffusion method. Front Vet Sci. 2025;12:1563208.
Crossref - Valgas C, Souza SM, Smânia EFA, Smânia A Jr. Screening methods to determine antibacterial activity of natural products. Braz J Microbiol. 2007;38(2):369-380.
Crossref - Balouiri M, Sadiki M, Ibnsouda SK. Methods for in vitro evaluating antimicrobial activity: A review. J Pharm Anal. 2016;6(2):71-79.
Crossref - Copeland LO, McDonald MB. Principles of seed science and technology. 5th ed. New York: Springer; 2001
- Younesikelaki FS, Ebrahimzadeh MH, Desfardi MK, Banala M, Marka R, Nanna RS. Optimization of seed surface sterilization method and in vitro seed germination in Althaea officinalis L. – An important medicinal herb. Indian J Sci Technol. 2016;9(28):1-6.
Crossref - Fu Y, Li X, Yuan X, et al. Alternaria alternata F3, a novel taxol-producing endophytic fungus isolated from the fruits of Taxus cuspidata: Isolation, characterization, taxol yield improvement, and antitumor activity. Appl Biochem Biotechnol. 2024;196(4):2246-2269.
Crossref - Chepkirui C, Stadler M. The genus Diaporthe: A rich source of diverse and bioactive etabolites. Mycol Prog. 2017;16:477-494.
- Gomes RR, Glienke C, Videira SIR, Lombard L, Groenewald JZ, Crous PW. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia. 2013;31:1-41.
- Wikee S, Lombard L, Crous PW, et al. Phyllosticta capitalensis, a widespread endophyte of plants. Fungal Divers. 2013;60(1):91-105.
Crossref - Grabka R, d’Entremont TW, Adams SJ, et al. Fungal endophytes and their role in agricultural plant protection against pests and pathogens. Plants. 2022;11:384.
- Swandiny GF, Filaila E, Priyanto JA, et al. Potent antibacterial and cytotoxic bioactive compounds from endophytic fungi Diaporthe sp. associated with Salacia intermedia. Arch Microbiol. 2025;207(2):40.
Crossref - Wikee S, Jaidee P, Wongkam S, McKenzie EHC, Hyde KD, Chukeatirote E. Antimicrobial activity of crude extracts of Phyllosticta spp. Mycology. 2013;4(2):112-117.
Crossref - Glienke C, Pereira OL, Stringari D, et al. Endophytic and pathogenic Phyllosticta species, with reference to those associated with Citrus Black Spot. Persoonia. 2011;26:47-56
- Cushnie TPT, Cushnie B, Lamb AJ. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int J Antimicrob Agents. 2014;44(5):377-386.
Crossref - Uwineza PA, Urbaniak M, Stepien L, Gramza-Michalowska A, Wa kiewicz A. Efficacy of Lamium album as a natural fungicide: Impact on seed germination, ergosterol, and mycotoxins in Fusarium culmorum-infected wheat seedlings. Front Microbiol. 2024;15:1363204.
Crossref - Wu F, Gao Y, Yang W, Sui N, Zhu J. Biological functions of strigolactones and their crosstalk with other phytohormones. Front Plant Sci. 2022;13:821563.
Crossref - Hejl AM, Koster KL. The allelochemical sorgoleone inhibits root H+-ATPase and water uptake. J Chem Ecol. 2004;30:2181-2191.
Crossref - Dos Reis JBA, Lorenzi AS, Vale HMM. Methods used for the study of endophytic fungi: a review on methodologies and challenges, and associated tips. Arch Microbiol. 2022;204(11):675.
Crossref - Hardoim PR, van Overbeek LS, Berg G, et al. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev. 2015;79:293-320.
Crossref - Toghueo RMK, Zabalgogeazcoa I, Pereira EC, Vazquez de Aldana BR. A Diaporthe fungal endophyte from a wild grass improves growth and salinity tolerance of Tritordeum and perennial ryegrass. Front Plant Sci. 2022;13:896755.
Crossref - Fite T, Kebede E, Tefera T, Bekeko Z. Endophytic fungi: Versatile partners for pest biocontrol, growth promotion, and climate change resilience in plants. Front Sustain Food Syst. 2023;7:1322861.
Crossref - Ameen M, Mahmood A, Sahkoor A, Zia MA, Ullah MS.The role of endophytes to combat abiotic stress in plants. Plant Stress. 2024;12:100435.
Crossref - Rodriguez RJ, White JF, Arnold AE, Redman RS. Fungal endophytes: Diversity and functional roles. New Phytol. 2009;182:314-330.
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.