


ISSN: 0973-7510
E-ISSN: 2581-690X
The profenofos (PFF), an organophosphate (OP) insecticide, is extensively use in agriculture to control lepidopteran pests and its indiscriminate use in agriculture causes accumulation of the pesticide and its residue in the ecosystem. Interestingly, the microorganisms play a crucial role in detoxification of pesticides and other xenobiotics compounds. One PFF degrading novel bacterium, Pseudomonas aeruginosa strain BRD1 was isolated through enrichment culture technique from an agricultural soil of Baharpur, Burdwan, Purba Bardhaman, West Bengal, India. The bacterium is rod shaped, motile and Gram-negative in nature and shown optimal growth in 100 ppm PFF at 32 °C. The isolate BRD1 was identified as Pseudomonas aeruginosa strain BRD1 using 16S rRNA gene based molecular phylogenetic approach, phenotypic characterization and whole genome analyses. Quantitative biodegradation analysis by High-performance liquid chromatography (HPLC) chromatograms reveals that the strain can degrade 76% of PFF after 96 hrs incubation. The HPLC and Electrospray Ionization Mass Spectrometry (ESI-MS) investigation indicates that the strain degrades the PFF via the formation of intermediate metabolites, 4-bromo-2-chlorophenol (BCP), 3-methoxyphenol, 2-chloro-4-benzoquinone, O-ethyl S-propyl phosphorothioic acid (EPPA), 3,4-dimethyl benzoic acid and phosphoric acid but, BCP was produced as a major hydrolytic intermediate. The genome of the bacterium possesses several OP and PFF pesticide degrading genes like bphD, glpD, estB_1, estB_2, pdeB, pdeN, pdeG, gstB_2, yfcF, gstB_3, cpg2, lap, pepN and encodes several OP degrading enzyme groups like phosphodiesterase, cytochrome p450, glutathione S-transferase, metal dependent hydrolase, metallophosphoesterase, amidohydrolase. The current study elucidates that strain BRD1 efficiently can remove PFF within a short time.
Profenofos, Pseudomonas aeruginosa strain BRD1, Biodegradation, BCP, HPLC, ESI-MS
Profenofos [(PFF), O-(4-bromo-2-chlorophenyl) O-ethyl S-propyl phosphorothioate] is an organophosphate insecticide and it acts on target insects as an acetylcholinesterase inhibitor.1 It was first registered in USA in 1982 to control pests.2 It was reported that PFF was commercially introduced as organophosphate (OP) insecticide when other OPs are banned by US EPA and European Commission.3 This broad-spectrum insecticide is extensively used in many countries like the United States, China, Pakistan, Australia, Japan, Brazil, Australia and India in various crop production like cotton, tobacco, chilli, etc.4-10
An important concern of the pesticide is that the PFF has wide ranges of toxicities and health hazardness to various non-target organisms. In plant system, it was reported that PFF can reduce the photosynthetic yield by inhibiting the photosystem I and II.11 In animal system, it was reported that PFF cause several developmental anomalies in Zebrafish.12 It has also been reported that indiscriminate and extensive application of PFF deteriorated the microbial community structure of soil as well as water and air ecosystem13,14 and ultimately collapsed the normal function of the ecosystem. At the last, the non-systematic and indiscriminate application of PFF may cause massive environmental issues in the near future.
In this circumstance, research work on promising microbial degradation of PFF from the environment is urgently required so that an ecosystem has been restored for a green globe. Several microbial cultures have been reported to profenofos biodegradation, these are as follows Stenotrophomonas sp. G115; Pseudomonas plecoglossicida PF1, P. aeruginosa PF2, and P. aeruginosa PF310; Pseudoxanthomonas suwonensis strain HNM16; Pseudomonas aeruginosa, Achromobacter xylosoxidans, Bacillus sp. and Citrobacter koseri17; Oceanobacillus iheyensis, Exiguobacterium profundum, Bacillus firmus18; Bacillus cereus, Aneurinibacillus migulanus19; Klebsiella oxytoca20; Pseudomonas luteola21; Rahnella sp. PFF222; Enterobacter cloacae MUG7523; Bacillus sp. PF124; Cupriavidus nantongensis X1T,25 Bacillus cereus strain PM38.26
During the course of review literature studies, we noticed that PFF may cause several hazardness to our environment in several ways and cause different health issues to the non-target organisms. To keep these problems in mind, the authors set the aims to isolate an efficient PFF degrading bacterial culture from the soil sample of Burdwan, West Bengal, India, for reporting the detailed investigation of microbial degradation of PFF for the first time from this part of the world.
Chemicals
Analytical standard grade profenofos (CAS No. 41198-08-7) with purity ≥ 95.0 % (HPLC area %) and 4-Bromo-2-chlorophenol (CAS No. 3964-56-5)with purity ≥98.5% (GC area %) were purchased from Sigma-Aldrich.
Collection of sample
The soil sample was collected aseptically from an agricultural field of Baharpur, Purba Bardhaman, West Bengal, India (latitude 23.259065 N and longitude 87.804315 E). The sample was taken in a sterile Whirl-pak bag (Sigma-Aldrich) and stored at 4 °C before use.
Media for enrichment
For isolation of PFF degrading bacterial cultures, enrichment culture technique has been carried out and profenofos (100 ppm) was supplemented as sole carbon source in the mineral salt media [(MSM, g/L: K2HPO4, 0.2; KH2PO4, 0.8; MgSO4·7H2O, 0.2; CaSO4, 0.1; (NH4)2SO4, 1.0; NaCl, 0.3; FeSO4·7H2O, 0.005; pH 7.2 ± 0.2) and supplemented with fotch trace solution (mg/L: MnSO4·H2O, 169; ZnSO4·7H2O, 288; CuSO4·5H2O, 250; NiSO4·6H2O, 26; CoSO4, 28; Na2MoO4·2H2O, 24; pH 7.2 ± 0.2)].27 One gram of soil sample was weighed and suspended in 25 ml of sterile MM broth media (supplemented with 100 ppm of PFF) and incubated for 7 days at 32 °C. Then, 1 ml of enriched suspension was taken and added in another fresh 25 ml of the same media supplemented with 100 ppm of PFF and incubated for 7 days at 32 °C. In this way after three transfers, the final enriched broth was serial diluted upto 10-6 and spread on fresh PFF (100 ppm) supplemented MM agar plate. After proper incubation at 32 °C, the isolated bacteria were obtained and sub-cultured on fresh same agar plate for 7 times after a 7-8 days interval to enhance the phenotypic stability. The stable bacteria were then purified in TSA media by 4- 5 times subculturing. The purified isolates were maintained in MM+PFF plate, TSA plate and TSA slant. The bacteria were preserved in 10% glycerol stock at -20 °C refrigerator for future investigation. Out of them, the BRD1 bacterium was chosen for further biodegradation study due to its fast PFF utilization potentiality.
Phenotypic characterization
The bacterium was subjected to various phenotypic characterization tests like Gram staining, KOH test, endospore staining, indole production, MR-VP test, phosphate solubilization, catalase, caseinase, amylase, lipase, gelatinase, citrate utilization, motility, DNA hydrolysis, phenylalanine deaminase, etc.28 The FE-SEM study was carried out according to the protocol.29,30
Identification of bacterial isolate BRD1
For phylogenetic identification of the bacterium, the genomic DNA was isolated according to the Murmur’s protocol.31 Amplification of 16S rRNA gene was carried out by an automated programmable thermal cycler (Bio-Rad) using two universal primers namely 27f (5’AGA GTT TGA TCC TGG CTC AG 3′) and 1492r (5’TAC GGT TAC CTT GTT ACG ACT T 3′). The amplified PCR product was sent to Biokart India Pvt. Ltd. for Sanger sequencing. The forward and reverse complement sequences were manually aligned and combined to produce a contig sequence. The contig sequence was studied both at the nucleotide BLAST tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome) in NCBI and 16S based identification at EzBioCloud server (https://www.ezbiocloud.net/identify). The species level of the bacterium was confirmed by ANI and DDH analysis of the whole genome through ANI tool of Ezbiocloud and TYGS, GGDC at Leibniz Institute DSMZ (https://ggdc-test.dsmz.de/home.php) respectively. The whole genome sequencing work was carried out from the service of Biokart India Pvt. Ltd. A phylogenetic tree was constructed through MEGA 11 software by bootstrap method employing 1000 bootstrap replications.
Biodegradation study
In order to investigate the 96 hrs biodegradation study, the isolate was grown in 25 ml broth for four separate media (24 hrs, 48 hrs, 72 hrs, 96 hrs) supplemented with 100 ppm PFF along with a control for each set. In course of extraction, the entire broth was sacrificed for centrifugation at 6000 g for 10 minutes. The cell free supernatant was collected and subjected for liquid-liquid extraction by adding equal volume of ethyl acetate (HPLC grade) and mixed well by vortexing. The upper ethyl acetate part was pipetted out in a fresh tube and dehydrated with anhydrous sodium sulphate (Na2SO4) and concentrated the sample by a rotary evaporator.32 After evaporation, 1 ml of HPLC grade acetonitrile was added to the dried sample, mixed properly followed by filtration by 0.22 µm syringe filter. The filtrate was used for estimation of parent compound quantitatively through HPLC (Chromaster, Hitachi Corporation, Japan) analysis.
Quantification of PFF degradation through HPLC Analysis
Different concentrations of standard profenofos solution (0, 50, 100, 150, 200 and 250 ppm) were prepared for standard curve preparation. In both cases of standard solutions and biodegradative samples, 20 µL of sample was loaded in RP-HPLC system with C18 reversed-phase column and UV detector at 250 nm. The mobile phase, acetonitrile: water (80:20) was applied with a flow rate of 1 ml/min. Quantification of residual profenofos was measured by comparing the area value from the standard curve and detection of major intermediates were carried out by comparing with RT values of their authentic standard.
Identification of PFF and intermediates through ESI-MS analysis
The Electrospray Ionization Mass Spectrometry (ESI-MS) direct infusion was carried out by LC-ESI-MS/MS of Waters Xevo® G2-XS QTof with a hybrid ADC detector. In the degradative sample, profenofos and BCP were primarily checked by comparing the m/z pattern with their authentic standards. For identification of the parent compound and their possible intermediates were examined by comparing their obtained m/z pattern with the mass spectrum of NIST, PubChem.
Identification of PFF and OP degrading genes and enzymes
The Prokka annotation of the chromosomal DNA of P. aeruginosa strain BRD1 was carried out and the OP and PFF gene list was prepared.33 The list of the chromosomal enzymes of P. aeruginosa strain BRD1 encoded for possible in OP and PFF degradation was prepared through NCBI prokaryotic genome annotation pipeline of submitted WGS data of the BRD1 strain.
Isolation and phenotypic characterization
The strain BRD1 was isolated through enrichment culture technique. In enhancement of phenotypic stability, the isolate was sub-cultured at a 7-8 days interval in PFF (100 ppm) supplemented MM agar (Figure 1a). The bacterium was purified after 5 times repeated streaking in fresh tryptone soya agar plates. Out of many, the BRD1 isolate was selected for its stable PFF utilizing characteristics on MM+PFF plates. The bacterium was maintained both in MM+PFF and TSA plate and TSA slant for further use of all experimental works.
The BRD1 isolate was a Gram-negative, rod shaped and motile in nature. The bacterium has a potentiality to solubilize phosphate and can utilize the citrate. The isolate is also potential to produce caseinase, gelatinase, catalase, DNase and lipase enzymes (Table 1). In FE-SEM studies, the bacterium was single rod in shape and had a length and breadth with 2.433-2.511 µm and 721.9-780.4 nm respectively (Figure 1b and 1c).
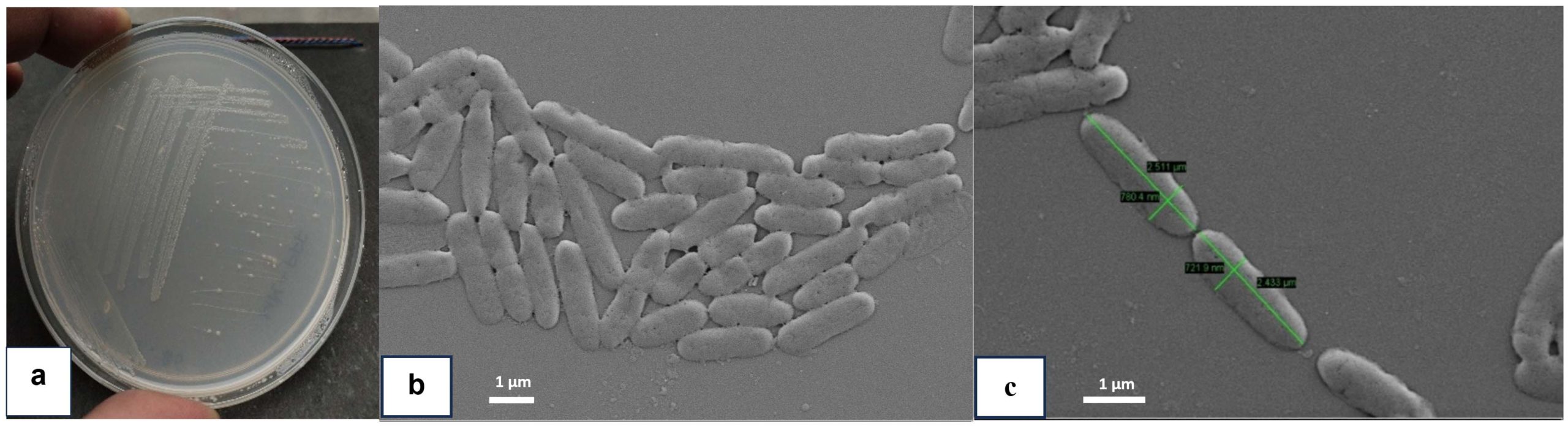
Figure 1. (a) BRD1 isolate in MM-PFF (100 ppm) Plate; (b, c) A fine structure of BRD1 isolate observed by FE-SEM
Table (1): Phenotypic Characterization of Pseudomonas aeruginosa strain BRD1
No. |
Tests |
Results |
|---|---|---|
1 |
Gram’s Staining |
– |
2 |
Color in Nutrient Broth |
Greenish |
3 |
Color in TSA plate |
Greyish |
4 |
Endospore Staining |
– |
5 |
Motility Test |
+ |
6 |
Phosphate Solubilization |
+ |
7 |
Citrate utilization |
+ |
8 |
Indole Production |
– |
9 |
Triple Sugar Iron Agar |
Red slant/ red butt |
10 |
Phenylalanine Deaminase production |
– |
11 |
KOH Test |
+ |
12 |
MR-VP test |
– |
13 |
Catalase production |
+ |
14 |
Caseinase production |
+ |
15 |
Lipase production |
+ |
16 |
Amylase production |
– |
17 |
DNase production |
+ |
18 |
Gelatinase production |
+ |
*‘+’indicates positive and ‘-’ indicates negative response in results
Phylogenetic identification
The genus level of the BRD1 isolate was identified as Pseudomonas by analysis of 16S rRNA gene based molecular phylogenetic approach followed by nucleotide BLAST at NCBI and 16S based identification at EZBioCloud. During pair-wise alignment, the 16S rRNA gene sequence of BRD1 isolate (16S GenBank accession PV649922) showed 100% sequence similarity with the type strain Pseudomonas aeruginosa strain JCM 5962 (Accession Number BAMA01000316) at EZBioCloud and 100% similarity with Pseudomonas aeruginosa strain 0201761-1 (Accession Number CP117300) of nucleotide BLAST in NCBI. The species level of BRD1 bacterium was confirmed as Pseudomonas aeruginosa by DDH, TYGS, ANI analysis of the whole genome sequences. The bacterium was identified as Pseudomonas aeruginosa strain BRD1 by analysing 16S rRNA gene, phenotypic characterizations, ANI, DDH and TYGS analyses. Therefore, the bacterial isolate was identified as Pseudomonas aeruginosa strain BRD1 as it shows 100% sequence similarity with closest phylogeny. One 16S rRNA gene based phylogenetic tree of BRD1 bacterium (Figure 2) was constructed with eight closely related strains, and one out group strain, Escherichia coli (ATCC 11775T). The 16S rRNA gene sequence of the BRD1 strain has been deposited at NCBI GenBank under accession number PV649922. The whole-genome shotgun work of the BRD1 strain has been deposited at NCBI Genome under accession number JBPJAS000000000 with version JBPJAS000000000.1. The associated metadata records of WGS are available under BioProject accession number PRJNA1276475 and the BioSample accession number is SAMN49073234.
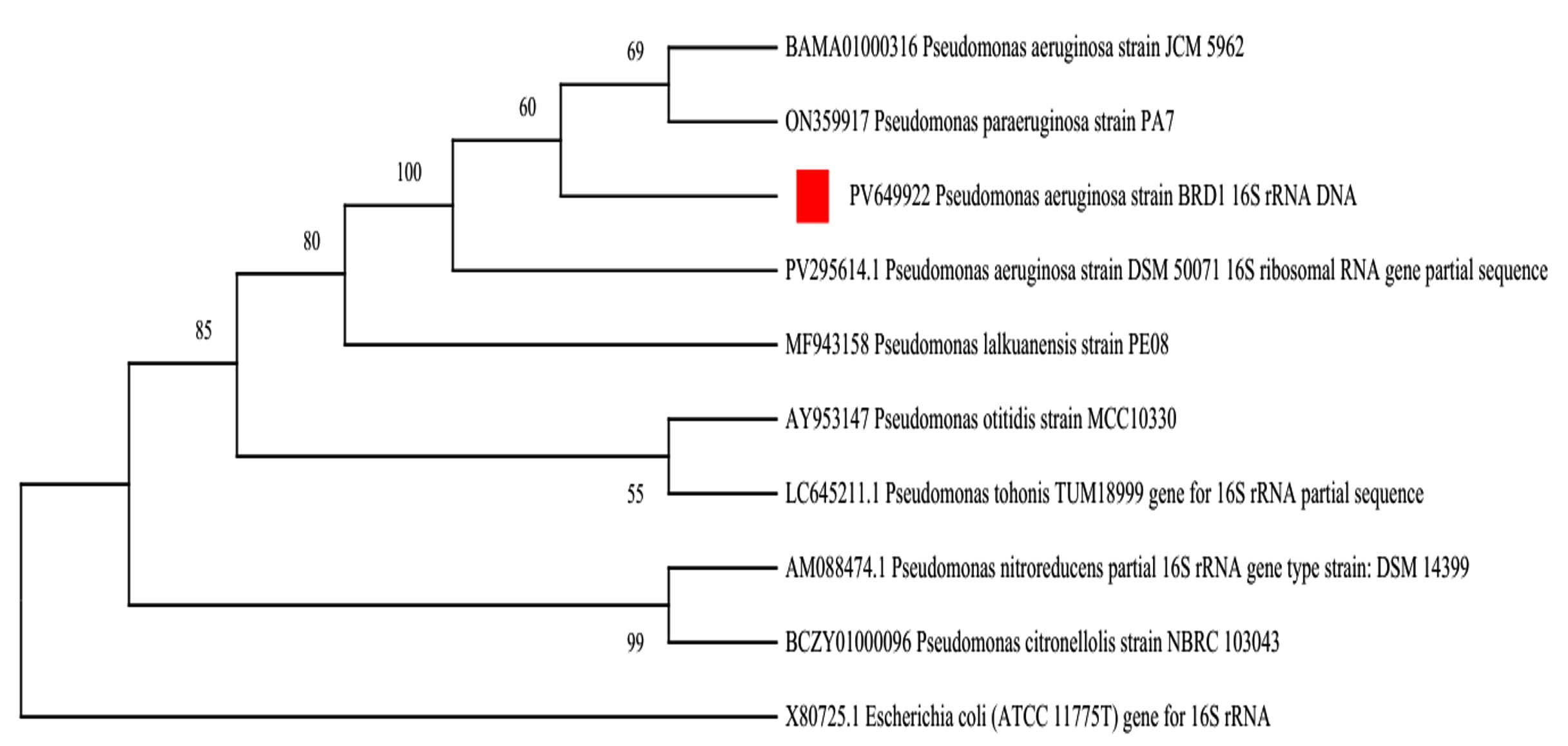
Figure 2. 16S rRNA gene based phylogenetic tree of Pseudomonas aeruginosa strain BRD1 with eight closely related phylogenetic groups and an outgroup strain Escherichia coli (ATCC 11775T) by bootstrap method, employing 1000 bootstrap replications
Biodegradation study
Quantification of PFF degradation through HPLC Analysis
In profenofos (PFF) biodegradation study, the analytical grade profenofos having RT value of 6.537 min (Figure 3a) and 4-Bromo-2-chlorophenol (BCP), major intermediate compound having RT of 4.760 min (Figure 3b). The detection and quantification of these compounds were carried out through HPLC analysis. BCP was detected in HPLC chromatograms of 24 hrs, 48 hrs, 72 hrs and 96 hrs samples (Figure 3c-3f). The quantitative estimation of residual PFF of the extracted samples were calculated through the standard curve. The detailed degradation study of PFF by Pseudomonas aeruginosa strain BRD1 was depicted in Figure 4 and it revealed that 46%, 62%, 65% and 76% PFF degradation was observed at each 24 h interval. The Figure 4 also represented the percentage of residual PFF as follows 54% (24 h), 38% (48 h), 35% (72 h) and 24% (96 h). During HPLC, the BCP was detected as a major intermediate and identified by comparing the RT value of the authentic standard. The degradation studies by HPLC analyses indicated that profenofos degraded through BCP as a major hydrolytic intermediate.
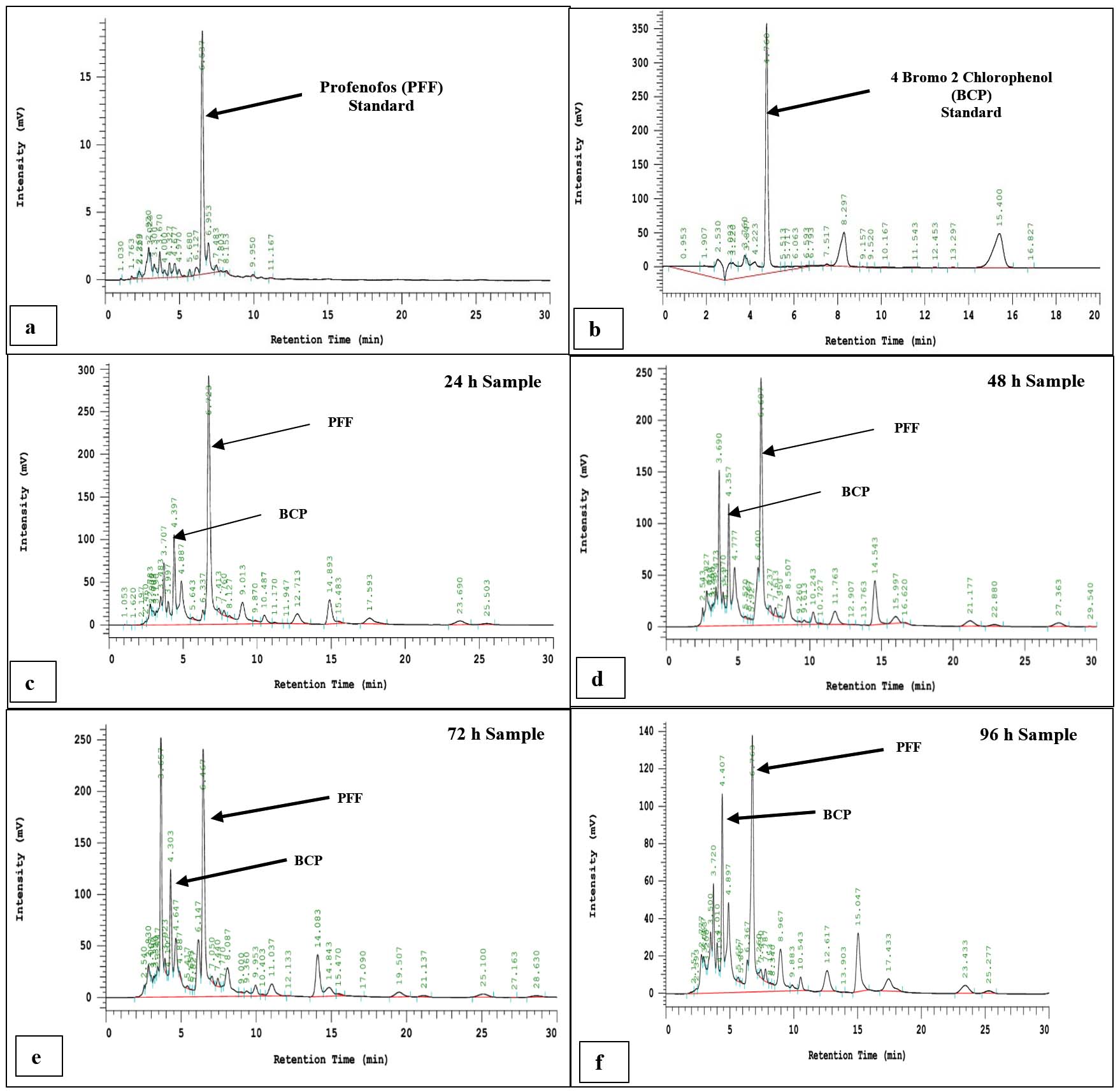
Figure 3. HPLC chromatogram of authentic standards and degradative samples of Pseudomonas aeruginosa BRD1 strain: (a) Profenofos (PFF) standard; (b) 4-Bromo-2-Chlorophenol (BCP) standard; (c) 24 hrs extracted sample; (d) 48 hrs extracted sample; (e) 72 hrs extracted sample, and (f) 96 hrs extracted sample
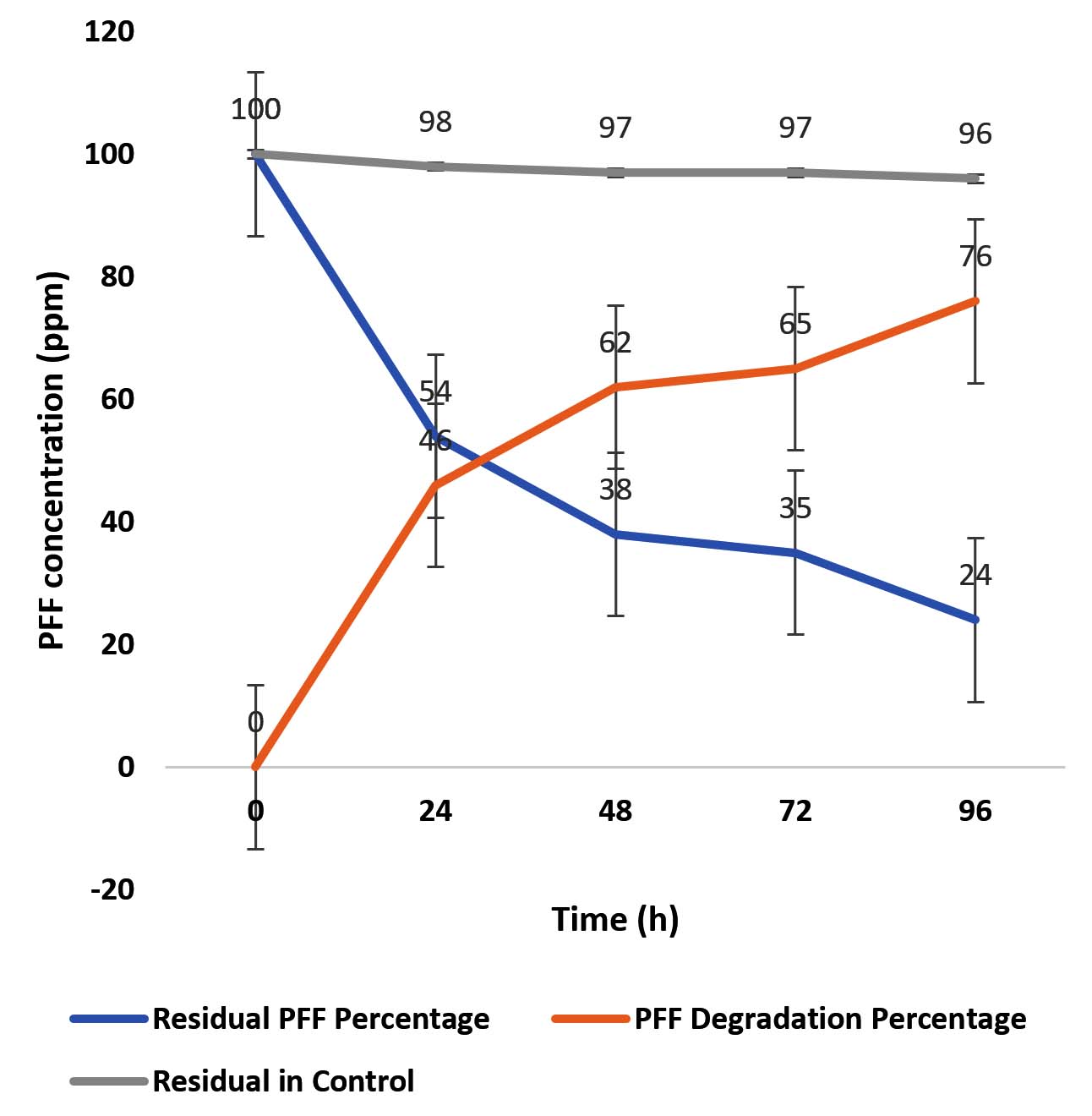
Figure 4. Residual and degradation percentage profile of profenofos after 24 hrs, 48 hrs, 72 hrs and 96 hrs samples by BRD1 strain
Identification of PFF and hydrolytic intermediates through ESI-MS analysis
In order to predict the identification of hydrolytic intermediates of parent compound PFF and construction of a possible degradation pathway by strain BRD1, the ESI-MS was performed. In ESI-MS analysis of the 72 h degradative sample, several compounds such as phosphoric acid (Figure 5a), 3-methoxyphenol (Figure 5b), 2-chloro-4-benzoquinone (Figure 5b), 3,4-dimethyl benzoic acid (Figure 5b), O-ethyl S-propyl phosphorothioic acid (Figure 5b), 4-bromo-2-chlorophenol (Figure 5c), some ion peaks of profenofos such as m/z 268.7874, 266.7913 and 297.2706 were detectable in Figure 5d and 302.7617, 344.7961 in Figure 5e and profenofos (Figure 5f) were detected and followed by identification of these intermediates compounds using both PubChem database and NIST Library search (Figure 5). The major ion peaks of profenofos (m/z 374.8225, 302.7617, 344.7961, 207.8278, 268.7874, 266.7913, 297.2706, 178.8901), BCP (m/z 207.8278, 206.8877, 208.8914, 169.8777, 127.5715, 63.3443), 2-chloro-4-benzoquinone (m/z 142.9502, 53.9544), 3-methoxyphenol (m/z 124.9496, 94.0855, 81.13, 67.2478), O-ethyl S-propyl phosphorothioic acid (m/z 184.9938, 110.9165, 109.004, 83.1187), 3,4-dimethyl benzoic acid (m/z 151.0418, 104.9122, 91.4585, 76.3300) and phosphoric acid (m/z 97.9118, 80.8224) were detected and matched respectively with mass pattern of the same compound respectively obtained from the NIST and PubChem database search. The HPLC analysis and ESI-MS analysis suggested that the strain Pseudomonas aeruginosa BRD1 can degrade the profenofos insecticide through a proposed possible degradation pathway (Figure 6).
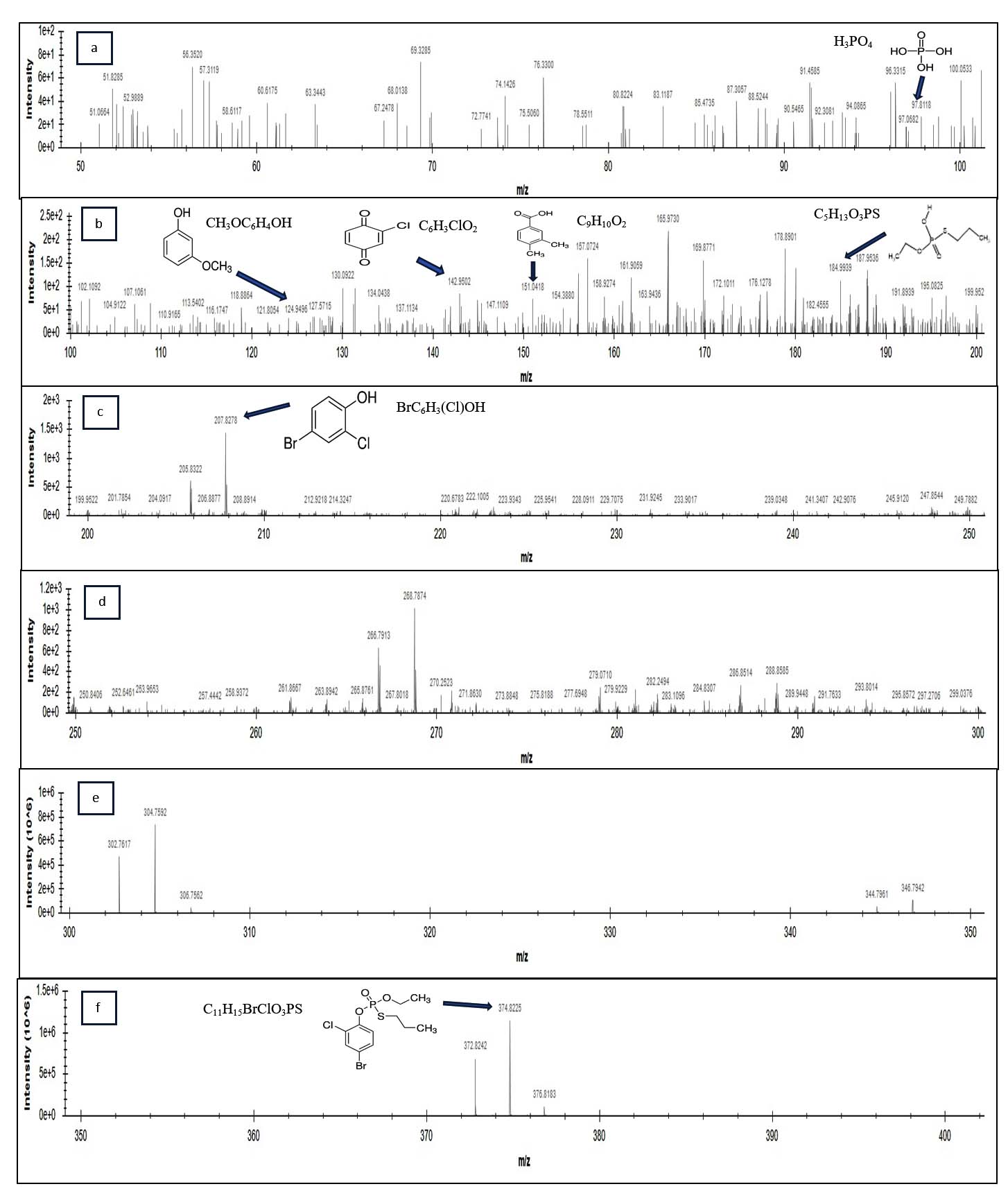
Figure 5. ESI-Mass Spectrometry analysis of 72 hrs biodegradative sample of Pseudomonas aeruginosa strain BRD1: (a) presence of phosphoric acid, (b) presence of 3-methoxyphenol, 2-chloro-4-benzoquinone, 3,4-dimethyl benzoic acid, and O-ethyl S-propyl phosphorothioic acid (EPPA) (c) presence of 4-bromo-2-chlorophenol (BCP) (d) m/z 268.7874, 266.7913 and 297.2706 ion peaks of PFF, (e) m/z 302.7617, 344.7961 ion peaks of PFF and (f) presence of parent compound Profenofos
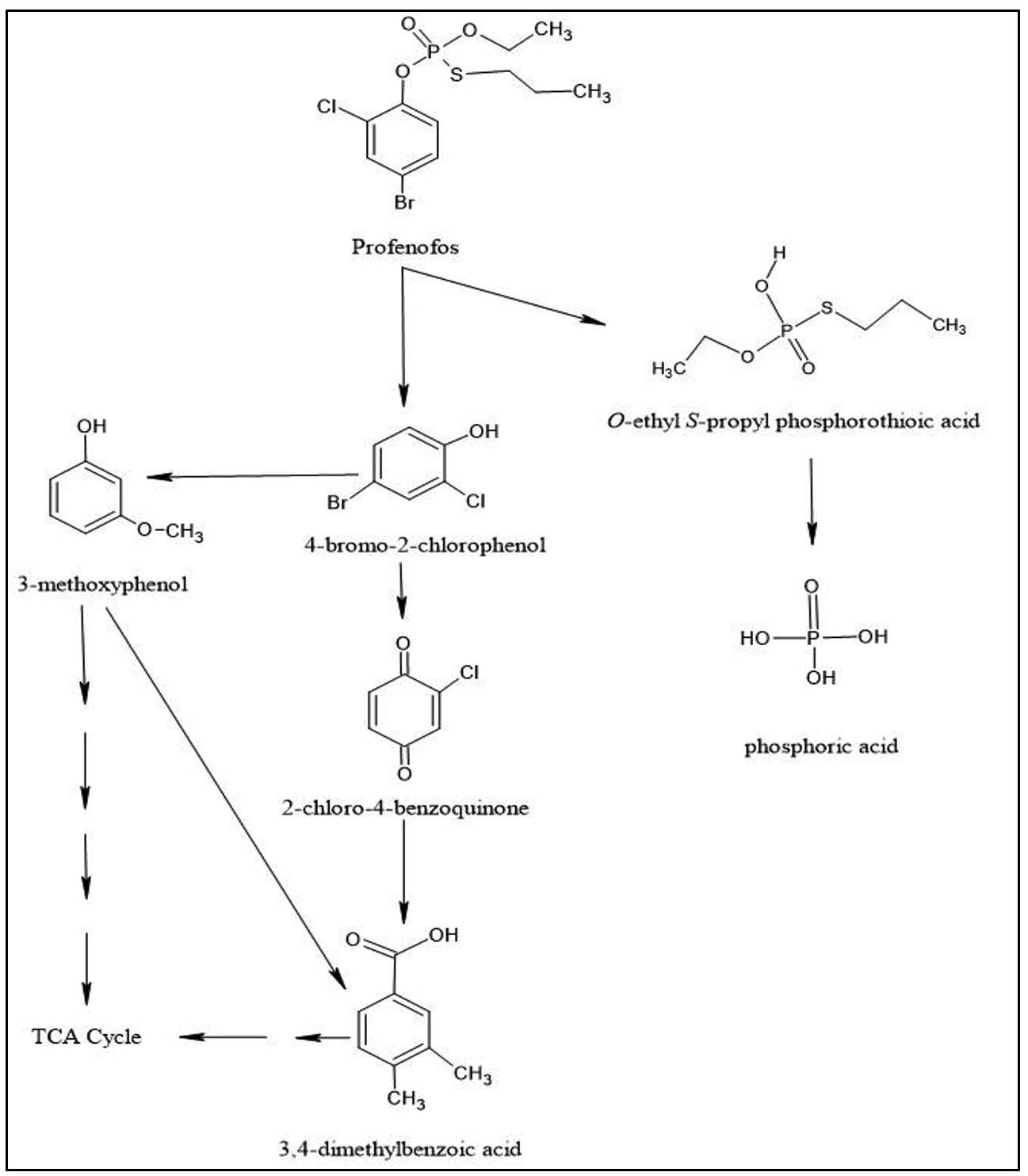
Figure 6. A possible pathway of profenofos biodegradation by Pseudomonas aeruginosa strain BRD1
PFF and OP degrading genes and enzymes
The Prokka annotation of the chromosomal DNA of Pseudomonas aeruginosa strain BRD1 shown several PFF and OP pesticide degrading genes such as bphD, glpD, estB_1, estB_2, pdeB, pdeN, pdeG, gstB_2, yfcF, gstB_3, cpg2, lap, pepN and those were listed in Table 2. On the other hand, chromosomal DNA of the bacterium encoded several OP groups enzymes like phosphodiesterase, cytochrome p450, glutathione s-transferase, metal dependent hydrolase, metallophosphoesterase, and amidohydrolase. The details of enzyme groups were mentioned in Table 3.
Table (2): List of OP and PFF degrading chromosomal genes of Pseudomonas aeruginosa strain BRD1 (Prokka annotation)
No. |
Name of gene |
Length (bp) |
Function |
|---|---|---|---|
1 |
bphD |
942 |
2-hydroxy-6-oxo-6-phenylhexa-2,4-dienoate hydrolase |
2 |
glpD |
1581 |
Aerobic glycerol-3-phosphate dehydrogenase |
3 |
estB_1 |
1176 |
Esterase EstB |
4 |
estB_2 |
648 |
Carboxylesterase 2 |
5 |
pdeB |
1596 |
putative cyclic di-GMP phosphodiesterase PdeB |
6 |
pdeN |
1578 |
putative cyclic di-GMP phosphodiesterase PdeN |
7 |
pdeG |
1581 |
putative cyclic di-GMP phosphodiesterase PdeG |
8 |
gstB_2 |
621 |
Glutathione S-transferase GstB |
9 |
yfcF |
636 |
Glutathione S-transferase YfcF |
10 |
gstB_3 |
624 |
Glutathione S-transferase GstB |
11 |
cpg2 |
1239 |
Carboxypeptidase G2 |
12 |
lap |
1611 |
Aminopeptidase |
13 |
pepN |
2658 |
Aminopeptidase N |
Table (3): List of the chromosomal enzymes of Pseudomonas aeruginosa strain BRD1 possible in OP and PFF degradation (NCBI PGAP)
| Group | Accession | Begin | End | Orientation | Name | Protein accession | Protein length | Locus tag |
|---|---|---|---|---|---|---|---|---|
| Phosphodiesterase | JBPJAS010000001.1 | 286961 | 289660 | minus | phosphodiesterase DipA | MGJ5993823.1 | 899 | ACSBR8_01300 |
| JBPJAS010000001.1 | 351171 | 351989 | plus | 3′,5′-cyclic-AMP phosphodiesterase | MGJ5993872.1 | 272 | ACSBR8_01545 | |
| JBPJAS010000001.1 | 546199 | 547134 | plus | glycerophosphodiester phosphodiesterase family protein | MGJ5994047.1 | 311 | ACSBR8_02440 | |
| JBPJAS010000001.1 | 556087 | 557268 | plus | cyclic di-GMP phosphodiesterase | MGJ5994058.1 | 393 | ACSBR8_02495 | |
| JBPJAS010000003.1 | 330821 | 333676 | plus | putative bifunctional diguanylate cyclase/phosphodiesterase | MGJ5994912.1 | 951 | ACSBR8_06830 | |
| JBPJAS010000004.1 | 253539 | 256133 | minus | bifunctional diguanylate cyclase/phosphodiesterase | MGJ5995215.1 | 864 | ACSBR8_08355 | |
| JBPJAS010000005.1 | 143835 | 144986 | minus | glycerophosphodiester phosphodiesterase | MGJ5995445.1 | 383 | ACSBR8_09515 | |
| JBPJAS010000005.1 | 147932 | 148405 | minus | metallophosphoesterase family protein | MGJ5995449.1 | 157 | ACSBR8_09535 | |
| JBPJAS010000006.1 | 86077 | 86385 | minus | Phosphodiesterase | MGJ5995655.1 | 102 | ACSBR8_10570 | |
| JBPJAS010000006.1 | 215129 | 215677 | minus | RNA 2′,3′-cyclic phosphodiesterase | MGJ5995780.1 | 182 | ACSBR8_11205 | |
| JBPJAS010000006.1 | 255279 | 256856 | plus | aminoglycoside response regulator cyclic di-GMP phosphodiesterase Arr | MGJ5995821.1 | 525 | ACSBR8_11435 | |
| JBPJAS010000007.1 | 61630 | 62202 | minus | ACP phosphodiesterase | MGJ5995923.1 | 190 | ACSBR8_11950 | |
| JBPJAS010000007.1 | 74762 | 76825 | plus | cyclic-di-GMP phosphodiesterase BifA | MGJ5995938.1 | 687 | ACSBR8_12025 | |
| JBPJAS010000019.1 | 47211 | 48338 | plus | glycerophosphodiester phosphodiesterase | MGJ5998033.1 | 375 | ACSBR8_22740 | |
| Metallophosphoesterase | JBPJAS010000022.1 | 25465 | 26208 | plus | Metallophosphoesterase | MGJ5998286.1 | 247 | ACSBR8_24015 |
| Amidohydrolase | JBPJAS010000002.1 | 214784 | 215818 | minus | amidohydrolase family protein | MGJ5994370.1 | 344 | ACSBR8_04095 |
| JBPJAS010000002.1 | 433041 | 434081 | minus | acetylpolyamine amidohydrolase AphA | MGJ5994574.1 | 346 | ACSBR8_05125 | |
| JBPJAS010000008.1 | 76861 | 78603 | plus | Amidohydrolase | MGJ5996167.1 | 580 | ACSBR8_13180 | |
| Metal dependent Hydrolase | JBPJAS010000011.1 | 175586 | 176473 | minus | metal-dependent hydrolase | MGJ5996918.1 | 295 | ACSBR8_17010 |
| JBPJAS010000023.1 | 30195 | 31280 | plus | MBL fold metallo-hydrolase | MGJ5998378.1 | 361 | ACSBR8_24475 | |
| JBPJAS010000030.1 | 57127 | 58365 | plus | M20/M25/M40 family metallo-hydrolase | MGJ5998937.1 | 412 | ACSBR8_27345 | |
| Cytochrome P450 | JBPJAS010000008.1 | 96872 | 98206 | minus | cytochrome P450 | MGJ5996185.1 | 444 | ACSBR8_13270 |
| JBPJAS010000011.1 | 162444 | 163700 | minus | cytochrome P450 family protein | MGJ5996912.1 | 418 | ACSBR8_16980 | |
| Glutathione S-transferase family | JBPJAS010000002.1 | 360575 | 361261 | plus | glutathione S-transferase family protein | MGJ5994507.1 | 228 | ACSBR8_04790 |
| JBPJAS010000005.1 | 280169 | 280789 | minus | glutathione S-transferase | MGJ5995567.1 | 206 | ACSBR8_10125 | |
| JBPJAS010000007.1 | 141868 | 142485 | minus | glutathione S-transferase N-terminal domain-containing protein | MGJ5995999.1 | 205 | ACSBR8_12340 | |
| JBPJAS010000016.1 | 4285 | 4986 | plus | glutathione S-transferase C-terminal domain-containing protein | MGJ5997644.1 | 233 | ACSBR8_20760 |
The bacterial isolate, BRD1 was identified as Pseudomonas aeruginosa strain BRD1 as it showed 100% sequence similarity with the type strains Pseudomonas aeruginosa strain JCM 5962 and Pseudomonas aeruginosa strain 0201761-1. The strain is motile in nature and forms greenish coloration in nutrient broth due to possible presence of water soluble pyocyanin and while the colonies appear as greyish in the TSA plate. It was reported that most of strains of Pseudomonas aeruginosa formed the pyocyanin pigment.34 The motile strains of Pseudomonas aeruginosa can detoxify the xenobiotics compounds of the environment through consortial degradation by their effective spreading and interaction with others microorganisms.35-37 Apart from this, the strain BRD1 is similar to other Pseudomonas aeruginosa species and potential to produce catalase, caseinase, gelatinase, lipase enzymes which have various biotechnological applications in the field of bioremediation, food processing, detergent, textile, pharmaceutical products, medical therapy, etc.38-40
The strain BRD1 degrades the profenofos as a sole carbon source and forms 4-bromo-2-chlorophenol, 2-chloro-4-benzoquinone, 3 methoxy phenol, O-ethyl S-propyl phosphorothioic acid, 3,4-dimethyl benzoic acid and phosphoric acid as major hydrolytic intermediates. The major metabolite, 4-bromo-2-chlorophenol is a less toxic compound than parent PFF.41 The previous report reveals that chlorophenol hampers the endocrine and reproductive health of fish.42 Chloro-benzoquinones can cause the oxidative DNA damage in E. coli cells along with the enhancement of intracellular ROS levels.43 3-methoxyphenol effects on various organisms is limited but, some methoxyphenols and their degradation products like guaiacol, ribavirin can be toxic to aquatic organisms when deposited into the aqueous environment.44,45 In PubChem comparative study, O-ethyl S-propyl phosphorothioic acid (CID 182032) is highly toxic due presence of sulphur and cause cholinesterase inhibition, 2-chloro-4-benzoquinone (CID 69671) is moderate in toxic for redox cycling and ROS generation, 4-bromo-2-chlorophenol (CID 19859) moderately toxic for membrane and protein, 3 methoxyphenol (CID 9007) cause oxidative stress and 3,4-dimethyl benzoic acid and phosphoric acid are less toxic to cells (https://pubchem.ncbi.nlm.nih.gov/). In accordance to literature survey, the chromosomal genome of the strain BRD1 has several PFF and OP pesticide degrading genes like bphD, glpD, estB_1, estB_2, pdeB, pdeN, pdeG, gstB_2, yfcF, gstB_3, cpg2, lap, pepN and several OP degrading enzyme groups like phosphodiesterase, cytochrome p450, glutathione s-transferase, metal dependent hydrolase, metallophosphoesterase, amidohydrolase.46,47
Authors went through a lot of research articles related to PFF degradation by various bacterial cultures to support the PFF degradation study by the strain BRD1. During this course of review literature, we observed several potential bacterial cultures that were reported for PFF biodegradation. It was reported that the bacterium, Stenotrophomonas sp. G1 can degrade 38% of PFF of 50 mg/L of initial concentration within 24 hr15 and Pseudomonas plecoglossicida PF1, P. aeruginosa PF2, and P. aeruginosa PF3 strains can utilize 95%, 93.1%, and 95.3% PFF respectively in 96 h with an initial concentration of 20 mg/L.10 A bacterial consortium including Pseudomonas aeruginosa, Achromobacter xylosoxidans, Bacillus sp. PF3, and Citrobacter koseri PF4 metabolized 100 mg/L PFF within 3 days and 200 mg/L within 8 days. But, the pure cultures of A. xylosoxidans PF1, P. aeruginosa PF2, Bacillus sp. PF3 and C. koseri PF4 can degrade 32%, 44%, 36% and 27% PFF respectively in 96 hrs of incubation.17
In the present research article, the bacterium Pseudomonas aeruginosa strain BRD1 can degrade 46% of PFF within 24 hr and 76% of PFF after 96 hrs alone. In comparison with other reported bacterial cultures, the strain BRD1 alone shows more efficiency in PFF degradation as a sole carbon source within a short passage of time. Elaborately, no other research article reported the PFF degradation by single bacterial culture except few reports.22,25 During PFF degradation study, 2-chloro-4-benzoquinone was detected and identified as another hydrolytic intermediate during ESI-MS analysis followed by PubChem library search which are less toxic than the parent compound. The proposed PFF utilization pathway (Figure 6) of the strain BRD1 indicates another route of conversion of 4-bromo-2-chlorophenol to 3,4-dimethyl benzoic acid through the formation of 2-chloro-4-benzoquinone and ultimately channelized into TCA cycle which is reported for the first time in the current article. The present article also reports the profenofos biodegradation by Pseudomonas aeruginosa strain BRD1 for the first time from the Purba Bardhaman district of West Bengal, India.
In this current investigation, the strain BRD1 can efficiently degrade 76% PFF insecticide of 100 µg/ml initial concentration within 96 hrs at mesophilic temperature and can detoxify PFF from the environment through bioremediation. The strain degrades PFF via the formation of 4-bromo-2-chlorophenol (BCP), 3-methoxyphenol, 2-chloro-4-benzoquinone, O-ethyl S-propyl phosphorothioic acid (EPPA), 3,4-dimethyl benzoic acid and phosphoric acid. Out of them, BCP was produced as a major hydrolytic intermediate. The strain possesses with several PFF and OPs insecticide degrading genes like bphD, glpD, estB_1, estB_2, pdeB, pdeN, pdeG, gstB_2, yfcF, gstB_3, cpg2, lap, pepN and enzyme groups like phosphodiesterase, cytochrome p450, glutathione s-transferase, metal dependent hydrolase, metallophosphoesterase, amidohydrolase. Apart from these, the bacterium is commercially important due to the presence of its phosphate solubilization, gelatinase, caseinase and lipase characteristics. It has been concluded that the strain Pseudomonas aeruginosa BRD1 is also a biotechnologically important microorganism and can be used in profenofos and other xenobiotic compounds degradation and industrial fields.
ACKNOWLEDGMENTS
The authors are thankful to the Department of Biotechnology, The University of Burdwan, West Bengal, for their support and for providing the platform to carry out the research work, and for providing the State Fund Fellowship. The authors are also grateful to the Bose Institute, Kolkata and Visva-Bharati University, Santiniketan, for ESI-MS and FE-SEM studies, respectively.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
SP conceptualized and supervised the study. SN performed data collection and experiments. SN and SP performed data analysis. SN and SP wrote, reviewed and revised the manuscript. Both authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
The 16S rRNA gene sequence of Pseudomonas aeruginosa strain BRD1 has been deposited at NCBI GenBank under Accession No. PV649922 and the whole-genome shotgun work have been deposited at NCBI Genome under Accession No. JBPJAS000000000 with version JBPJAS000000000.1. The associated metadata for WGS is available under BioProject Accession No. PRJNA1276475, BioSample Accession No. SAMN49073234 and SRA Accession No. SRR34337497.
ETHICS STATEMENT
Not applicable.
- Kushwaha M, Verma S, Chatterjee S. Profenofos, an acetylcholinesterase inhibiting organophosphorus pesticide: A short review of its usage, toxicity, and biodegradation. J Environ Qual. 2016;45(5):1478-1489.
Crossref - US Environmental Protection Agency. Interim Reregistration Eligibility Decision (IRED): Oxamyl. US Environmental Protection Agency, Office of Prevention, Pesticides, and Toxic Substances; 2000. Publication EPA-738-R-00-015
- Gotoh M, Sakata M, Endo T, Hayashi H, Seno H, Suzuki O. Profenofos metabolites in human poisoning. Forensic Sci Int. 2001;116(2-3):221-226.
Crossref - Radwan MA, Abu-Elamayem MM, Shiboob MH, Abdel-Aal A. Residual behaviour of profenofos on some field-grown vegetables and its removal using various washing solutions and household processing. Food Chem Toxicol. 2005;43(4):553-557.
Crossref - Li X, Li S, Liu S, Zhu G. Lethal effect and in vivo genotoxicity of profenofos to Chinese native amphibian (Rana spinosa) tadpoles. Arch Environ Contam Toxicol. 2010;59(3):478-483.
Crossref - Swarnam TP, Velmurugan A. Pesticide residues in vegetable samples from the Andaman Islands, India. Environ Monit Assess. 2013;185(7):6119-6127.
Crossref - Van Toan P, Sebesvari Z, Blasing M, Rosendahl I, Renaud FG. Pesticide management and their residues in sediments and surface and drinking water in the Mekong Delta, Vietnam. Sci Total Environ. 2013;452-453:28-39.
Crossref - Rajesh L, David M. Hepatotoxic potentials of profenofos on wistar albino rats: a histopathological study. Int J Pharm Biol Arch. 2014;5(1):70-75
- Mishra AK, Chaudhary S, Kumar A, Chandrasekharan H. Identification of Simple Sequence Repeats in chloroplast and mitochondrial genome of wheat. In: International Conference on Computing for Sustainable Global Development (INDIACom) 2014:265-270.
Crossref - Siripattanakul-Ratpukdi S, Vangnai A S, Sangthean P, Singkibut S. Profenofos insecticide degradation by novel microbial consortium and isolates enriched from contaminated chili farm soil. Environ Sci Pollut Res. 2015;22(1):320-328.
Crossref - Sidhu GK, Singh S, Kumar V, Dhanjal DS, Datta S, Singh J. Toxicity, monitoring and biodegradation of organophosphate pesticides: a review. Crit Rev Environ Sci Technol. 2019;49(13):1135-1187.
Crossref - Pamanji R, Yashwanth B, Bethu MS, Leelavathi S, Ravinder K, Rao JV. Toxicity effects of profenofos on embryonic and larval development of Zebrafish (Danio rerio). Environ Toxicol Pharmacol. 2015;39(2):887-897.
Crossref - Gomez F, Martinez-Toledo MV, Salmeron V, Rodelas B, Gonzalez-Lopez J. Influence of the insecticides profenofos and diazinon on the microbial activities of Azospirillum brasilense. Chemosphere.1999;39(6):945-957.
Crossref - Gupta A, Singh UB, Sahu PK, et al. Linking soil microbial diversity to modern agriculture practices: a review. Int J Environ Res Public Health. 2022;19(5):3141.
Crossref - Deng S, Chen Y, Wang D, et al. Rapid biodegradation of organophosphorus pesticides by Stenotrophomonas sp. G1. J Hazard Mater. 2015;297:17-24.
Crossref - Talwar MP, Ninnekar HZ. Biodegradation of pesticide profenofos by the free and immobilized cells of Pseudoxanthomonas suwonensis strain HNM. J Basic Microbiol. 2015;55(9):1094-1103.
Crossref - Jabeen H, Iqbal S, Anwar S, Parales R E. Optimization of profenofos degradation by a novel bacterial consortium PBAC using response surface methodology. Int Biodeterior Biodegradation. 2015;100:89-97.
Crossref - Isworo S, Purwanto, Sabdono A. Identification of bacteria indigenous selected Rawa Pening lake owns best degradation capability of the organophosphate pesticides (malathion and profenofos). Asian J Appl Sci. 2016;9(2):41-52.
Crossref - Palanimanickam A, Sepperumal U. Degradation of Profenofos in Soil inoculated with Bacillus cereus and Aneurinibacillus migulanus. Indian J Appl Microbiol. 2018;21(3):35-42.
Crossref - Gonzales-Condori EG, Ramirez-Revilla SA, Villanueva-Salas JA. Role of Eisenia foetida in the degradation of profenofos in presence of native bacterial communities. Rev Mex Ing Quim. 2020;19(Sup. 1) 45-57.
Crossref - Putra IWWP, Setiyo Y, Gunam IBW, Anggreni AAMD. Isolation and identification of profenofos pesticide degrading bacterium from soil sample of Bedugul, Indonesia. IOP Conf Ser: Earth Environ Sci. 2021;724(1):12-37.
Crossref - Verma S, Chatterjee S. Biodegradation of profenofos, an acetylcholine esterase inhibitor by a psychrotolerant strain Rahnella sp. PFF2 and degradation pathway analysis. Int Biodeterior Biodegradation. 2021;158:105-169.
Crossref - Ghani MU, Asghar HN, Niaz A, Ahmad Zahir Z, Nawaz MF, Haggblom MM. Efficacy of rhizobacteria for degradation of profenofos and improvement in tomato growth. Int J Phytoremediation. 2022;24(5):463-473.
Crossref - Mahajan R, Verma S, Chatterjee S. Biodegradation of organophosphorus pesticide profenofos by the bacterium Bacillus sp. PF1 and elucidation of initial degradation pathway. Environ Technol. 2023;44(4):492-500.
Crossref - Fang L, Zhou Y, Chen T, et al. Efficient biodegradation characteristics and detoxification pathway of organophosphorus insecticide profenofos via Cupriavidus nantongensis X1T and enzyme OpdB. Sci Total Environ. 2023;862:160782.
Crossref - Pakar NP, Rehman FU, Mehmood S, et al. Microbial detoxification of chlorpyrifos, profenofos, monocrotophos, and dimethoate by a multifaceted rhizospheric Bacillus cereus strain PM38 and its potential for the growth promotion in cotton. Environ Sci Pollut Res Int. 2024;31(27):39714-39734.
Crossref - Malghani S, Chatterjee N, Hu X, Zejiao L. Isolation and characterization of a profenofos degrading bacterium. Journal of Environmental Sciences. 2009;21(11):1591-1597.
Crossref - Leboffe MJ, Pierce BE. Microbiology: Laboratory Theory and Application. 3rd ed. Morton Publishing Company; Englewood, Colorado, USA. 2010
- Hayat MA. Principles and Techniques of Electron Microscopy: Biological Applications. Vol 3. Van Nostrand Reinhold Co; 1970.
- Das R, Goswami S, Pal M, et al. Assessment of antibacterial activity of leaf extracts of Crinum asiaticum L. against five gram negative fish pathogenic bacterial strains. Microbe. 2025;7:100319.
Crossref - Kamlage B. Methods for General and Molecular Bacteriology. Edited by P. Gerhardt, R. G. E. Murray, W. A. Wood and N. R. Krieg. 791 pages, numerous figures and tables. American Society for Microbiology, Washington, D.C., 1994. Food / Nahrung. 1996;40(2):103.
Crossref - Pailan S, Saha P. Chemotaxis and degradation of organophosphate compound by a novel moderately thermo-halo tolerant Pseudomonas sp. strain BUR11: evidence for possible existence of two pathways for degradation. Peer J. 2015;3:e1378.
Crossref - The Galaxy Community. The Galaxy platform for accessible, reproducible, and collaborative data analyses: 2024 update. Nucleic Acids Res. 2024;52(W1):W83-W94.
Crossref - Saleem H, Mazhar S, Syed Q, Javed MQ, Adnan A. Bio-characterization of food grade pyocyanin bio-pigment extracted from chromogenic Pseudomonas species found in Pakistani native flora. Arab J Chem. 2021;14(3):103005.
Crossref - Pandey G, Jain RK. Bacterial chemotaxis toward environmental pollutants: role in bioremediation. Appl Environ Microbiol. 2002;68(12):5789-5795.
Crossref - Qattan SYA. Harnessing bacterial consortia for effective bioremediation: targeted removal of heavy metals, hydrocarbons, and persistent pollutants. Environ Sci Eur. 2025;37(1):85.
Crossref - Bhatt P, Bhatt K, Huang Y, Li J, Wu S, Chen S. Biofilm formation in xenobiotic-degrading microorganisms. Crit Rev Biotechnol. 2023;43(8):1129-1149.
Crossref - Hana MM, Abdeen EE. Pseudomonas aeruginosa: Prevalence in Human and Animal Products, Virulence and Antibiotic Resistance Aspects. Alex J Vet Sci. 2024;82:21-35
Crossref - Zambare V, Nilegaonkar S, Kanekar P. A novel extracellular protease from Pseudomonas aeruginosa MCM B-327: enzyme production and its partial characterization. N Biotechnol. 2011;28(2):173-181.
Crossref - Mokrani S, Nabti EH. Recent status in production, biotechnological applications, commercial aspects, and future prospects of microbial enzymes: A comprehensive review. Int J Agric Sci Food Technol. 2024;10(1):006-020.
Crossref - Dadson OA, Ellison CA, Singleton ST, et al. Metabolism of profenofos to 4-bromo-2-chlorophenol, specific and sensitive exposure biomarker. Toxicology. 2013;306:35-39.
Crossref - Ge T, Han J, Qi Y, et al. The toxic effects of chlorophenols and associated mechanisms in fish. Aquat Toxicol. 2017;184:78-93.
Crossref - Chen Z, Zhou Q, Zou D, et al. Chloro-benzoquinones cause oxidative DNA damage through iron-mediated ROS production in Escherichia coli. Chemosphere. 2015;135:379-386.
Crossref - An Z, Sun J, Han D, et al. Theoretical study on the mechanisms, kinetics and ecotoxicity assessment of OH-initiated reactions of guaiacol in atmosphere and wastewater. Sci Total Environ. 2019;685:729-740.
Crossref - Jiang J, An Z, Li M, et al. Comparison of ribavirin degradation in the UV/H2O2 and UV/PDS systems: Reaction mechanism, operational parameter and toxicity evaluation. J Environ Chem Eng. 2023;11(1):109193.
Crossref - Raj A, Kumar A, Khare PK. The looming threat of profenofos organophosphate and microbes in action for their sustainable degradation. Environ Sci Pollut Res Int. 2024;31(10):14367-14387.
Crossref - Pailan S, Sengupta K, Saha P. Microbial metabolism of organophosphates: key for developing smart bioremediation process of next generation. In: Arora P, ed. Microbial Technology for Health and Environment. Springer Nature Singapore; 2020:349-373. Microorganisms for Sustainability; vol 22.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.