


ISSN: 0973-7510
E-ISSN: 2581-690X
The objective of this study was to identify and study the effect of colchicine treatment on the growth and enzyme production of Pycnoporus coccineus KKUPN1. Molecular identification using universal primer ITS1 and ITS4 found that KKUPN1 was Pycnoporus coccineus (ID: KU202741). The effect of colchicine when culturing P. coccineus KKUPN1 in Czapex doc broth and potato dextrose broth (PDB) containing 0, 0.05 and 0.1% (v/v) colchicine for 7 and 14 d was studied. The result showed that in the early stage all treatments containing colchicine gave better growth than the treatments without colchicine. However, 5 d after inoculation, the growth rates were not significantly different in all treatments. The colony morphology of P. coccineus KKUPN1 isolated from Czapex doc broth containing colchicine showed the production of orange-red pigment that was not present from the medium without colchicine. In PDB containing 0.05 and 0.1% (v/v) colchicine for 7 d, the colonies had orange-red pigment from the edge of the colony to the center, while 0% (v/v) colchicine produced sparse mycelia with slight pigment. The enzyme test of P. coccineus KKUPN1 cultured in Czapex doc broth or PDB with colchicine added produced results that showed they did not produce a hemicellulolytic enzyme and a lignin modification enzyme, but could produce a cellulolytic enzyme that was detected by CMC agar and esculin agar but could not be detected by cellulose agar.
Colchicine, Enzyme production, Pycnoporus coccineus.
Pycnoporus coccineus (Scarlet Bracket) is a cosmopolitan group of white rot fungi belonging to the kingdom Fungi, division Basidiomycota, class Agaricomycetes, order Polyporales, family Polyparaceae. Scarlet Bracket is one of the most common and colorful brackets that can be found even in dry weather growing on sticks and wood. The fruiting-body is orange scarlet, fan shaped, firm bracket attaching along the straight edge to the wood. It can be solitary but more common in large groups on sticks and logs, and refreshed after rain. The white-rot fungi of the genus Pycnoporus are red polypores, divided into four species: Pycnoporus cinnabarinus, native to the North Temperate Zone; Pycnoporus coccineus, which occurs mostly in countries bordering the Indian and Pacific Oceans; Pycnoporus sanguineus, found in tropical and subtropical regions of the northern and southern hemispheres; and Pycnoporus puniceus in Africa and India1, 2. In Thailand there are 10 species of Pycnoporus3. P. sanguineus has been intensively studied because of its metabolites production and the production of several extracellular enzymes belonging to the category of oxyredutases that are known to be mainly responsible for the degradation process of lignocellulosic waste. The fungus produces an important secondary metabolite that is cinnabarin4. White rot fungi are good lignin degraders and have the potential to be used in industry5. Industrially important extracellular enzymes from filamentous fungi, such as laccases, are often N-glycosylated with high mannose structures. In nature, the role of these laccases is to degrade lignin to gain access to the other carbohydrates in wood (cellulose and hemicellulose)6. Vegetal biomass is composed of three main polymers: lignin, cellulose and hemicelluloses. Cellulose is organized as glucose polymeric long lineal chains, arranged in different levels of fibrils that are associated by intramolecular interactions. Hemicelluloses, in contrast, are ramified polymers composed of different types of carbohydrates and phenolic compounds, among which, xylan, glucan and arabinoglucan form the sugar backbone of the structure7.
Colchicine is an effective chemical mutagen. It binds to tubulin and prevents its polymerization into microtubules8. Colchicine inhibits the formation of spindle fibers and effectively arrests mitosis at the anaphase stage. At this point, the chromosomes have multiplied but cell division has not yet taken place resulting in polyploidy cells. Colchicine treatment is the classical method of inducing the doubling of the chromosome number9. This was studied to enhance the degrading ability of the microcrystalline cellulose in the mycelia of Lentinula edodes by incubating the mycelia in an autopolyploidization medium containing colchicine for more than one week at a low temperature. The results showed that almost all of the microcrystalline cellulose in the agar medium was degraded by the colchicine treated strain10. Autodiploid strains were induced by colchicine treatment of Aspergillus niger WU-2223L, a citric acid producing strain. In shaking culture, the autodiploid strain yielded higher citric acid than the parental strain11. The objective of this study was to identify and study the effect of colchicine on the growth and enzyme production of Pycnoporus coccineus KKUPN1.
Microorganisms and Identification
P. coccineus KKUPN1 was obtain from the Fungal and Mushroom Laboratory, Plant Pathology Section, Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University. For identification, P. coccineus was incubated on potato dextrose agar (PDA) at 28ºC for 7 d, after which a plug (0.7 cm in diameter) of the fungal mycelium was transferred to 50 ml of potato dextrose broth (PDB) in a 250 ml Erlenmeyer flask. Cultures were grown at 28ºC with shaking at 120 rpm for 7 d. The mycelium was filtered from the liquid medium through filter paper (Whatman No. 1) and kept at -20 ºC for 24 hr before DNA extraction. Total DNA extraction was prepared by a modified method of Lee et al12 and Zang et al13 For amplification of the ITS1-5.8S-ITS2 regions, genomic DNA were amplified using universal primers ITS1 (52 -TCCGTAGGTGAACCTGCGG-32 ) and ITS4 (52 -TCCTCCGCTTATTGAT ATGC-32 )14. The PCR reaction mixture and the PCR thermal cycle were according to the protocol of Sutthisa et al15. Amplification was performed in 50 µl of reaction mixture containing 100 ng genomic DNA, 2.0 µM of each primer, 0.25 units Taq polymerase, 2.5 mM MgCl2, 0.2 mM of each dNTP (dATP, dCTP, dGTP and dTTP), 10 µl 5X PCR buffer and sterile distilled water to a final volume of 50 µl. Amplifications were performed in a Gradient DNA Thermal Cycler programmed with the following parameters: 95°C for 5 min; followed by 30 cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for 1 min; and a final incubation at 72°C for 5 min. Amplification products (5 µl of a 50 µl reaction) were electrophoresed in 2% agarose gels with TBE running buffer, stained with ethidium bromide and either scanned into a computer imaging file or photographed16. PCR products were sequenced by First BASE Laboratories Sdn Bhd.
Colchicine Treatment
P. coccineus KKUNN01 grown on PDA at 28ºC for 7 d was transferred into PDB and Czapex doc broth medium containing 0, 0.05 and 0.1% (v/v) colchicine solution in test tubes and incubated at 28ºC. After 7 and 14 d the mycelium were transferred to PDA in 9 cm diameter petri dishes and the mycelia growth were observed and colony diameters were measured.
Mycelia Stain
Mycelia mats were stained with Giemsa stain by flooding absolute methanol to cover the mycelia mat for 3 min and then drying. Then the slides were dipped in working Giemsa stain for 30 min, after staining the mycelia mat was rinsed with distilled water, left to dry and photographed under a microscope (Zeiss Primo Star). Measurements of the width of the mycelium and arthrospores (50 sporres) were observed.
Enzyme Production
The detection of the enzyme production was according to the methods of Pointing17 and Abdel-Raheem and Shearer18, including:
Cellulolytic enzyme assays: Three methods were used to detect the cellulolytic enzymes, including cellulose agar clearance, dye staining of carboxymethylcellulose agar and esculin plus iron agar.
Hemicellulolytic (xylanolytic) enzyme assays: Dye staining of the xylan agar was used to detect the hemicellulolytic enzyme.
Lignin modifying enzyme assays: Staining after growth on lignin agar was used to detect the lignin modifying enzyme.
Identification
The amplification of the genomic DNA of P. coccineus KKUPN1 by universal primers ITS1 and ITS4 produced a single amplification product of approximately 591 bp. When comparing the nucleotide sequences to the GenBank database it revealed a similarity with Pycnoporus coccineus strain IUM0049 (GenBank ID: KP255840.1) with 100% identity (Figure 1), and the sequencing files are stored in GenBank under the accession number KU202741.1.
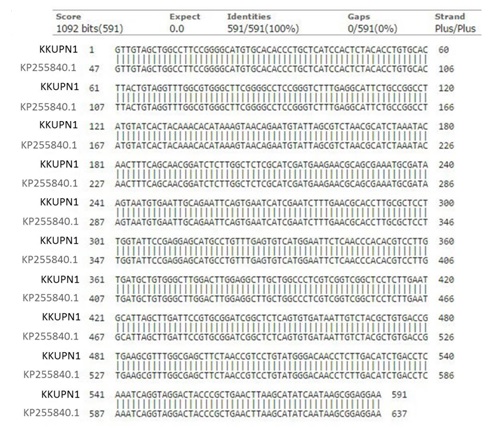
Figure 1. Sequence alignment of Pycnoporus coccineus KKUPN1 (KU202741) and KP255840.119.
Colchicine Treatment
The mycelial growth rate of P. coccineus KKUPN1 after culture in Czapex doc broth and PDB medium containing colchicine for 7 and 14 d showed that during the early stage of mycelial growth the growth rates were slightly different. The treatment with added colchicine tended to give growth that was greater than without colchicine. Then 5 d after inoculation, for the mycelial growth rate there were no significant differences between all the treatments (Table 1).It has been reported that the nuclei of Pleurotus ostreatus increased in diameter after treatment with colchicine solution and P. ostreatus yields were increased by up to 82% compared to the control. The stem and the diameter of the basidiocarp also enlarged in the colchicine-treated P. ostreatus [20].
The colony morphology of P. coccineus KKUPN1 isolated from the Czapex doc broth containing colchicine showed the production of an orange-red pigment in contrast to the medium without colchicine. The growth of the mycelia radiated from the center of the colony. During the early stage, P. coccineus KKUPN1 produced a powder-like mycelia. Subsequent phases developed orange-red pigment in circles as the colony grew. P. coccineus KKUPN1 cultured in PDB medium containing 0.05 and 1% colchicine for 7d showed a lot of orange-red pigment from the edge of the colony to the center. The control treatment (0% colchicine) produced sparse mycelia with only a little pigment. Therefore, when cultured in the PDB medium containing 0 and 0.05% colchicine for 14d it produced sparse mycelia with a little pigment, while the 0.1% colchicine produced more orange-red pigment. However, it was still less than in the 7d cultures (Figure 2). A previous report indicated that the colony diameter of P. coccineus (Fr.) Bod. and Sing. was about 7.5cm 2w after inoculation on PDA. The colony contained four morphological areas: powder-like, cottony, synematoue and pore-fruiting with bright orange pigments [21]. The mycelial growth and colony characteristics of Pycnoporus sanguineus grown on PDA medium were observed 7d after inoculation at 25±1ºC. The colony grew moderately rapid and usually covered the 9cm petri dish in 4d. The mycelia were white at first, slightly raised, thin and translucent, with some appressed opaque white areas. Color began to appear after 3 to 4d, as granules of moderate orange over the inoculum [22]. The colony morphology of Pycnoporus cinnabarinus on YM medium at 20°C after 13d was white to cinnamon, flat, velutinous or powdery. The hyphae were hyaline or pigmented, monomitic and with simple septate. Chlamydospores were intercalary and ovoid. Arthrospores were cylindrical and hyaline.
Table (1):
Growth rates of Pycnoporus coccineus KKUPN1 after induction by colchicine.
| Colchicine treatment | Colony diameter (cm) | ||||
|---|---|---|---|---|---|
| Days after incubation | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Czapex dox broth | |||||
| 0% 7days | 0.69±0.22 b | 2.31±0.24 c | 4.19±0.28 c | 6.53±0.28ab | 8.50±0.00 ns |
| 0.05% 7days | 0.75±0.24 b | 2.32±0.16 c | 4.47±0.18 bc | 6.51±0.27 ab | 8.50±0.00 ns |
| 0.1% 7days | 0.81±0.15 ab | 2.60±0.12 ab | 4.57±0.20 b | 6.61±0.22 ab | 8.50±0.00 ns |
| 0% 14 days | 0.73±0.25 b | 2.49±0.17 bc | 4.28±0.13 c | 6.35±0.07 b | 8.50±0.00 ns |
| 0.05% 14 days | 0.83±0.21ab | 2.58±0.14 ab | 4.71±0.20 ab | 6.76±0.21 a | 8.50±0.00 ns |
| 0.1% 14 days | 1.03±0.12 a | 2.78±0.04 a | 4.86±0.10 a | 6.81±0.02 a | 8.50±0.00 ns |
| CV (%) | 22.07 | 6.19 | 4.23 | 3.07 | 0.00 |
| PDB | |||||
| 0% 7 days | 1.19±0.16 a | 2.42±0.19ab | 4.48±0.13 b | 7.53±0.26 b | 8.50±0.00 ns |
| 0.05% 7 days | 1.08±0.33 ab | 2.28±0.38 ab | 4.48±0.54 b | 7.58±0.63 b | 8.50±0.00 ns |
| 0.1% 7 days | 1.04±0.01 ab | 2.05±0.06 b | 4.14±0.12 b | 7.12±0.20 b | 8.50±0.00 ns |
| 0% 14 days | 0.75±0.34bc | 3.07±0.64 a | 6.12±1.23 a | 8.50±0.00 a | 8.50±0.00 ns |
| 0.05% 14 days | 0.42±0.07 c | 2.20±0.62ab | 5.52±1.08 ab | 8.25±0.61 a | 8.50±0.00 ns |
| 0.1% 14 days | 0.90±0.50 ab | 2.42±1.11 ab | 6.75±2.07 a | 8.50±0.00 a | 8.50±0.00 ns |
| CV (%) | 32.44 | 25.24 | 20.99 | 4.85 | 0.00 |
Note: Different superscripts within the same column indicate a significant difference (P < 0.05).
Table (2):
Width of Pycnoporus coccineus KKUPN1 mycelium and arthrospores after induction with colchicine.
| Colchicine treatment | Mycelium diameter (µ) | Arthrospore (µ) | ||
|---|---|---|---|---|
| Czapexdox broth | PDB | Czapexdox broth | PDB | |
| 0% 7days | 2.10±0.52 c | 3.30±1.06 bc | 1.5-2.5 x 2.5-5.0 | 1.5-2.0 x 3.0-5.0 |
| 0.05% 7days | 2.95±0.60 ab | 4.25±1.32 ab | 1.5-2.0 x 3.0-5.0 | 2.0-2.5 x 3.0-6.0 |
| 0.1% 7days | 2.95±0.50 ab | 4.35±0.88 a | 1.0-3.0 x 3.0-5.0 | 2.0-3.0 x 3.0-6.0 |
| 0% 14 days | 2.50±0.67 bc | 3.05±0.64 c | 1.5-2.0 x 3.0-4.5 | 1.0-2.0 x 3.0-5.5 |
| 0.05% 14 days | 3.45±1.26 a | 3.75±0.72 abc | 1.5-2.0 x 3.5-6.0 | 2.0-2.5 x 3.0-6.0 |
| 0.1% 14 d | 3.55±0.44 a | 4.10±0.61 ab | 1.5-2.0 x 3.5-6.0 | 2.0-3.0 x 3.0-6.0 |
| CV (%) | 24.60 | 23.89 | ||
Note: Different superscripts within the same column indicate a significant difference (P < 0.05).
The P. coccineus KKUPN1 cultured in Czapex doc broth or PDB containing colchicine showed that the mycelial of P. coccineus KKUPN1 was wider than when grown without colchicine. The mycelia of P. coccineus KKUPN1 cultured in PDB medium containing colchicine was wider than that cultured in Czapex doc broth medium containing colchicine at both 7 and 14 d (Table 2; Figure 3). It was also found that it produced short rod arthrospores that were not different in all the treatments. The arthrospore sizes varied from 1.0-3.0 x 2.5-6.0 μ in the Czapex doc broth containing colchicine and from 1.0-3.0 x 3.0-6.0 μ in PDB containing colchicine (Table 2; Figure 4). The advancing hyphae at the colony periphery of P. coccineus were 2.0-2.5 µ in diameter with frequent clamp connections. The surface hyphae of the powder-like areas were 1.5-2.2 µin diameter, frequently branched and septate [21]. Those in the culture media were from single cell arthrospores [23].

Figure 2. Colony morphology of Pycnoporus coccineus KKUPN1 on PDA after induction with colchicine treatment in Czapexdox broth and PDB medium. A-F in Czapexdox broth and G-L in PDB.
A, G: 0% colchicine 7 d;
B, H: 0.05% colchicine 7 d;
C, I: 0.1% colchicine 7 d;
D, J: 0% colchicine 14 d;
E, K: 0.05% colchicine 14 d;
F, L: 0.1% colchicine 14 d.
Enzyme production
The enzyme activity test for P. coccineus KKUPN1 culture in Czapex doc broth or PDB containing colchicine had results showing that P. coccineus KKUPN1 did not produced a hemicellulolytic enzyme via dye staining of xylan agar. A positive reaction indicated the degradation of the substrate by endoxylanase and β-xylosidase. Lignin enzyme production was determined to be negative when using lignin agar. This assay can be useful in determining the ability of a fungus to utilize a lignin substrate. The method indicates the degradation of phenolic components in lignin [17]. The cellulolytic enzyme can be detected by CMC agar and esculin agar but it isnegative in cellulose agar because the clearance of the cellulose within the growth media is difficult to assess. The CMCase activity of P. sanguineus was higher than the activity towards crystalline cellulose [7].
The CMC agar test showed that a high concentration of colchicine and long exposure to colchicine resulted in the high production of cellulolytic enzymes in both the Czapex doc broth and PDB medium (Table 3; Figure 5). CMC is a substrate for endoglucanase and can be used as a test for endoglucanase and β-glucosidase activity [24].
The esculin agar test showed that with a concentration of 0.1% colchicine for 7 d in both media, P. coccineus KKUPN1 could produce the most cellulolytic enzyme with a black color in the medium. While the other treatment showed no difference, except in the control treatment (0% colchicine) at 7 and 14 d in the PDB, which could produce a minimal amount of enzyme (Table 3; Figure 6). The hydrolysis of cellobiose to glucose is achieved by β-glucosidase. This enzyme is probably ubiquitous among cellulolytic fungi producing hydrolytic endoglucanase or cellobiohydrolase. The activity of β-glucosidase can be detected by the growth of the test fungi on agar containing esculin as the sole carbon source. The splitting of the substrate by the enzyme yields glucose and a coumarin product that reacts with iron sulphate to produce a black color in the growth medium [17, 18].
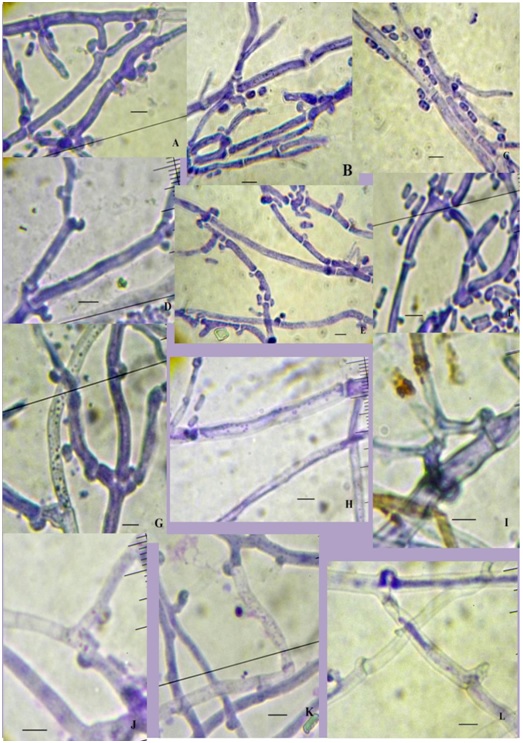
Figure 3. Pycnoporus coccineus KKUPN1 mycelia. A-F in Czapexdox broth and G-L in PDB.
A,G: 0% colchicine 7 d;
B, H: 0.05% colchicine 7 d;
C, I: 0.1% colchicine 7 d;
D, J: 0% colchicine 14 d;
E, K: 0.05% colchicine 14 d; F,
L: 0.1% colchicine 14d. ( = 5 µ)
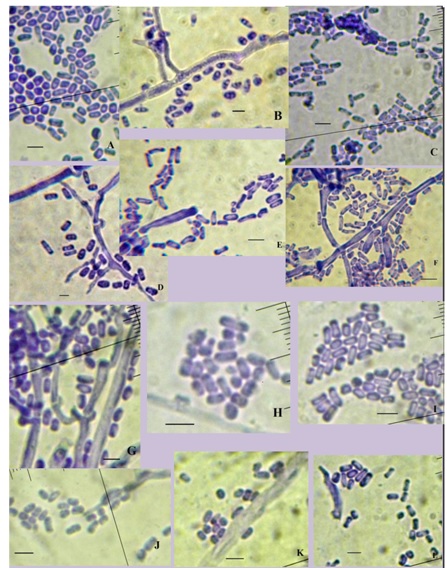
Figure 4. Arhrospores of Pycnoporus coccineus KKUPN1. A-F in Czapexdox broth and G-L in PDB.
A,G: 0% colchicine 7 d;
B, H: 0.05% colchicine 7 d;
C, I: 0.1% colchicine 7d;
D, J: 0% colchicine 14 d;
E, K: 0.05% colchicine 14 d;
F, L: 0.1% colchicine 14 d.( ←= 5 µ)
Mutagens enhanced the enzyme production of Pycnoporus cinnabarinus. Colchicine and ethidium bromide showed increases in enzyme production with increased concentrations [25]. White rot fungi have the capacity to exploit all wood components due to the secretion of a variety of lignocellulolytic enzymes [26]. White rot fungi are good lignin degraders and have the potential to be used in industry. Phellinus sp., Daedalea sp., Trametesversicolor and Pycnoporus coccineushave been selected due to their relatively high ligninolytic enzyme activity. Cellulase enzyme assays showed no significant cellulose activity detected in the enzyme preparations of T. versicolor and Phellinus sp. This low cellulolytic activity further suggests that these two white rot strains are of more interest in lignin degradation [5]. The cellulolytic properties of two white rot fungi, Bjerkanderaadustaand Pycnoporussanguineus, cultivated on wheat straw agar medium, were characterized and compared. Enzymes from P. sanguineuswere more robust as they better resisted a1 h incubation at high temperatures (up to 80ºC), and exhibiting activity and stability in a pH range from 2 to 8. B. adustaand P. sanguineus when grown on a natural cellulosic substrate (wheat straw) had a number of cellulolytic activities with different characteristics [7]. Pretreatment of cellulosic substrates with white-rot fungi achieved high levels of saccharification of the holocellulose [27].
Table (3):
Enzyme production of Pycnoporus coccineus after induction with colchicine.
| Colchicine treatment | Hemicellulolytic enzyme | Lignin modify enzyme | Cellulolytic enzyme | ||
|---|---|---|---|---|---|
| Xylan agar | Lignin agar | Cellulose agar | CMC agar | Esculin agar | |
| Czapecdox broth | |||||
| 0% 7 days | – | – | – | 0.33±0.06 c1/ | ++ |
| 0.05% 7 days | – | – | – | 0.53±0.06 b | ++ |
| 0.1% 7 days | – | – | – | 0.60±0.10 ab | +++ |
| 0% 14 days | – | – | – | 0.33±0.06 c | ++ |
| 0.05% 14 days | – | – | – | 0.63±0.12 ab | ++ |
| 0.1% 14 days | – | – | – | 0.67±0.06 a | ++ |
| CV (%) | 21.13 | ||||
| PDB | |||||
| 0% 7 days | – | – | – | 0.37±0.06 cd | + |
| 0.05% 7 days | – | – | – | 0.50±0.10 bc | ++ |
| 0.1% 7 days | – | – | – | 0.60±0.17 ab | +++ |
| 0% 14 days | – | – | – | 0.30±0.00 d | + |
| 0.05% 14 days | – | – | – | 0.67±0.15 a | ++ |
| 0.1% 14 days | – | – | – | 0.63±0.06 ab | ++ |
| CV (%) | 21.13 | ||||
Note: Different superscripts within the same column indicate a significant difference (P < 0.05).
– = noreaction, + = slight reaction, ++ = moderate reaction, +++ = strong reaction
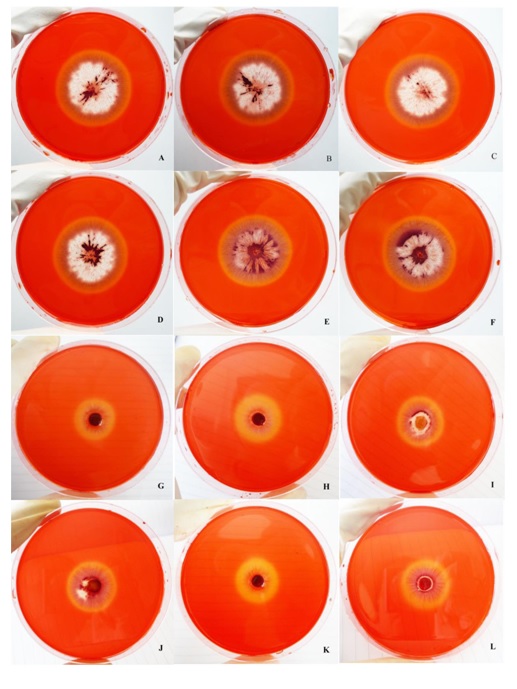
Figure 5.Cellulolytic enzyme assay by dye stained of carboxymethylcellulose agar of Pycnoporus coccineus KKUPN1. A-F in Czapex dox broth And G-L in PDB.
A, G: 0% colchicine 7 d;
B, H: 0.05% colchicine 7 d;
C, I: 0.1% colchicine 7 d;
D, J: 0% colchicine 14 d;
E, K: 0.05% colchicine 14 d;
F, L: 0.1% colchicine 14 d.
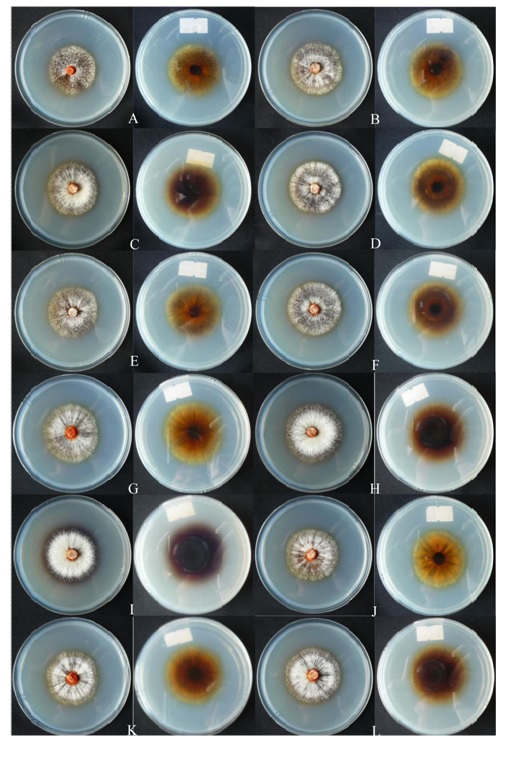
Figure 6.Cellulolytic enzyme assay by esculin plus iron agar of Pycnoporus coccineus KKUPN1. A-F in Czapex dox broth and G-L in PDB.
A, G: 0% colchicine 7 d;
B, H: 0.05% colchicine7 d;
C, I: 0.1% colchicine 7 d;
D, J: 0% colchicine 14 d;
E, K: 0.05% colchicine 14 d;
F, L: 0.1% colchicine 14 d
The work studied the inoculation of P. coccineus KKUPN1 (ID: KU202741) in media containing colchicine at different concentrations and for different durations. We found that at the early stage all treatments that contained colchicine tended to grow better than without colchicine, but after that the growth rates were not significantly different. Colchicine effected the pigmentation and mycelial width. In our studies we found that P. coccineus KKUPN1 did not produce a hemicellulolytic enzyme or a lignin modifying enzyme, but could produce a cellulolytic enzyme that was detected by CMC agar and esculin agar but could not be detected by cellulose agar. However, more study is necessary.
ACKNOWLEDGMENTS
Grateful acknowledgement is made of the Holistic Watershed Management Cluster of Khon Kaen University for providing financial support. Thanks for the use of research equipment from the Agricultural Research Center for Sustainable Economy, Khon Kaen University and Center of Excellence on Agricultural Biotechnology (AG-BIO/PERDO-CHE). We would like to thank Dr. Jolyon Dodgson from Faculty of Science, Mahasarakham University for proofreading.
- Uzan, E., Nousiainen, P., Balland, B., Sipila, J., Piumi, F., Navarvo, D., Asther, M., Record E., Lomascolo, A. High redox potential laccases from the ligninolytic fungi Pycnoporus coccineus and Pycnoporus sanguineus suitable for white biotechnology: from gene cloning to enzyme characterization and application. J. Appl. Microbiol., 2009, 108: 2199-2213.
- Ryvarden, L., Johansen, I. Pycnoporus Karst”, in “A preliminary Polypore Flora of East Africa. (Ed. L. Ryvarden), Oslo, Norway, Fungiflora, 1980.
- Thongkred, P., Lotrakul, P., Prasongsuk, S., Imai, T., Punnapayak, H. Pxidation of polycyclic aromatic hydrocarbons by a tropical isolate of Pycnoporus coccineus and its laccase. Science Asia, 2011, 37: 225-233.
- Baumer, J.D., Mas Diego, S.M., Pacheco, S.M.V., Morgago, A.F.M., Furigo, A.F. Jr. Comparative study of mycelial growth and production of cinnabarin by different strains of Pycnoporus sanguineus. BioFar, 2008, 2: 1-15.
- Liew, C.Y., Husaini, A., Hussain, H., Muid, S., Liew, K.C., Roslan, H.A. Lignin biodegradation and ligninolytic enzyme studies durig biopilping of Acacia mangium wood chips by tropical white rot fungi. World J. Microbiol. Biotechnol., 2011, 27: 1457-68.
- Vite-Vallejo, O., Palomares, L.A., Dantan-Gonzalez, E., Ayala-Castro, H.G., Martinez-Anaya, C., Valderrama, B., Folch-mallol, J. The role of N-glycosylation on the enzymatic activity of Pycnoporus sanguineus laccase. Enzyme Microb. Technol., 2009, 45: 233-239.
- Quiroz-Castaneda, R.E., Balcazar-Lopez, E., Dantan-Gonzalez, E., Martinez, A., Folch-Mallol, J., Martinez-Anaya, C. Characterization of cellulolytic activities of Bjerkandera adusta and Pycnoporus sanguineus on solid wheat straw medium. Electronic J. Biotechnol., 2009, 12: 1-8.
- Brown, T.A. Mutations are Also Caused by Chemical and Physical Mutagens. 2nd Edn., Oxford, Wiley-Liss, 2002.
- Wongpiyasatid, A., Hormchan, P., Rattanadilok, N. Preliminary test of polyploidy induction in cotton (Gossypium arboretum) using colchicine treatment. Nat. Sci., 2003, 37: 27-32.
- Toyama, H. Enhancement of microcrystalline cellulose degrading ability in Lentinula edodes by autopolyploidization at reduced temperature. Intl. J. of Adv. Biotec. and Res., 2003: 4: 308-314.
- Sarangbin, S., Morikawa, S., Kirimura, K., Usami, S. Formation of autodiploid strains in Aspergillus niger and their application to citric production from starch. J. Ferment. Bioeng., 1994: 77: 474-478.
- Lee, Y.M., Yong-Keel, C., Byung-Re, M. PCR-RFLP and sequence analysis of the rDNA ITS region in the Fusarium spp. J. of Microb., 2000, 38(2): 66-73
- Zang, Z., Zhang, J., Wang, Y., Zheng. Molecular detection of Fusarium oxysporum f.sp. niveum and Mycosphaerella melonis in infected plant tissues and soil. FEMS Microb. Letters, 2005, 249: 39-47
- White, T.J., Bruns, T., Lee S., Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. in PCR Protocols: A Guide to Methods and Applications (ED. M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White), Academic Press, Inc., New York, 1990, pp. 315-322.
- Sutthisa, W., sanoamuang, N., Chuprayoon, S. Evaluation of amplified rDNA restriction analysis (ARDRA) for the identification of Fusarium species, the causal agent associated with mulberry root rot disease in Northeastern Thailand. J. Agric. Technol., 2010; 6: 379-390.
- Sambrook, J., Fritsch, E.F., Maniatis, T. Molecular cloning, a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989.
- Pointing, S.B. Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal Divers., 1999: 2: 17-33.
- Abdel-Raheem, A., Shearer, C.A. Extracellular enzyme production by freshwater ascomycetes. Fungal Divers., 2002: 11: 1-19.
- Park, J.W., Kang, H.W., Ha, B.S., Kim, S.I., Kim, S., Ro, H.S. Strain-dependent response to Cu2+ in the expression of laccase in Pycnoporus coccineus. Arch. Microbiol., 2015, In press.
- Hosseini, S.Z., Karimzadeh, G., Mohammadi, G.E. Effect of colchicine on nuclei diameter, yield and some morphological characteristics of oyster mushroom (Pleurotuse ostreatus). J. Agric. Sci., 2009: 19: 253-261.
- Lin, W., Hollingsworth, P.J., Volz, P.A. Morphology of Pycnoporus coccineus (Fr.) Bond. and Sing in vitro. Taiwania, 1982: 27: 39-47.
- Albert, S., Pandya, B., Padhiar, A. Evaluation of colony characteristics and enzymatic activity of some fungi for potential use in co-culture for bio pulping. Asian J. Biol. Life Sci., 2012: 1: 83-89.
- Watanabe, T. Pictorial Atlas of Soil and Seed Fungi, Morphologies of Cultures Fungi and Key to Species. 3rd Edn., CRC Press, Tayler & Francis Group Boca Raton, London, Newyork, 2010, pp. 141.
- Buswell, J.A., Cai, Y.J., Chang, S.T., Peberdy, J.F., Fu, S.Y., Yu, H.S. Lignovellulolytic enzyme profiles of edible mushroom fungi. World J. Microbiol. Biotechnol, 1996: 12: 537-542.
- Khanam, R., Prasuna, R.G. Strain improvement of white rot fungi Pycnoporus cinnabarinus with the influence of physical and chemical mutagens for enhancing laccases production. J. Sci. Industr. Res., 2014: 73: 331-337.
- Martinez, A.T., Speranza, M., Ruiz-Duenas, F.J., Ferreira, P., Camarero, S., Guillen, F., Martinez, M.J., Gutierrez, A., Del-Rjo, J.C. Biodegradation of lignocellulosics microbial, chemical, and enzymatic aspects of the fungal attack of lignin. International Microbiol., 2005: 8: 195-204.
- Lee, J.W., Gwak, K.S., Park, J.Y., Park, M.J., Choi, D.H., Kwon, M., Choi, I.G. Biological pretreatment of softwood Pinus densiflora by three white rot fungi. J. Microbiol., 2007: 45: 485-491.
© The Author(s) 2017. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.