


ISSN: 0973-7510
E-ISSN: 2581-690X
Limited soil-soluble Phosphorus and inefficient application of chemical phosphate fertiliser cause a phosphorus (P) deficit, reducing crop yield. In this study, healthy soybean (Glycine max) plants from the Shirala region of Maharashtra, India, were used to isolate endophytic and rhizospheric bacteria. These bacteria were evaluated for phosphate-solubilising capacity and other PGP attributes. Ten bacterial isolates showed clear phosphate solubilisation halos on Katznelson’s and Bose agar medium, with the isolate SWK having the highest solubilisation index (3.50). Compared with the uninoculated control, a quantitative study in Pikovskaya’s liquid medium showed a 704.7% increase in soluble Phosphorus. The isolate SWK exhibited several important plant growth–promoting characteristics, including measurable nitrogenase activity (4.86 ± 0.32 µmol C2H4 per hour), positive ACC deaminase function, substantial formation of indole-3-acetic acid (28.7 ± 1.8 µg mL-1 with L-tryptophan supplied) and effective siderophore secretion, reflected by a D/d ratio of 2.18. Physiological and biochemical analyses further revealed that the isolate SWK is a Gram-negative, bacillus-shaped bacterium possessing considerable metabolic versatility, enabling it to adapt efficiently to diverse environmental conditions. The 16S rRNA method revealed that the isolate was Enterobacter hormaechei strain SWK (LC923381). The impact of bacterial inoculation on soybean growth was investigated in pot culture using different inoculation methods. An ANOVA showed substantial variation in plant height, stem thickness, root length, and total biomass among treatments (P < 0.001). The isolate SWK applied via combined rhizosphere irrigation and foliar spray produced the highest growth responses, including maximum plant height (46.8 ± 2.2 cm) and biomass (13.8 ± 0.7 g plant-1). Overall, Enterobacter hormaechei strain SWK showed superior plant growth–promoting efficiency, highlighting its potential as a biofertiliser for sustainable soybean cultivation.
Isolation, Rhizosphere, Morphological, Soybean
Phosphorus is key to numerous plant biochemical and physiological activities, such as energy metabolism, nucleic acid synthesis, membrane synthesis, and cellular signalling. However, its availability in soil is often limited due to rapid conversion into insoluble forms, which restricts efficient absorption by plants. Worldwide, almost 52.3 billion tonnes of phosphorus-containing fertilisers are applied each year to maintain the soil–plant phosphorus balance and help overcome this limitation.1 However, low fertiliser use efficiency and rising production costs occur because crops absorb only 15%-20% of applied Phosphorus, with the remainder becoming immobilised through fixation processes.2
The ongoing depletion of Phosphorus, a non-renewable resource, threatens global food security and sustainable agricultural development.3,4 Phosphorus deficiency severely restricts plant growth and productivity, particularly in subtropical regions and highly weathered soils.5-7 Consequently, the emergence of environmentally clean alternatives to fertilisers with chemicals, along with strategies to improve phosphorus availability and utilization efficiency, has become a critical priority in modern agricultural systems.
Phosphate-solubilising bacteria (PSB) represent an important group of beneficial microorganisms capable of releasing chemical compounds (acids), chelators, & enzymes that transform insoluble phosphorus into forms accessible for plant uptake.8,9 Many PSBs not only solubilise Phosphorus but also fix nitrogen, produce phytohormones, synthesise siderophores, and have enzyme activity that improves stress tolerance, all contributing to increased plant growth and productivity. Thus, PSBs have been extensively explored as biofertilisers to reduce chemical fertiliser use, increase soil fertility, and reduce pollution.10-12
Despite their potential, the field performance of phosphate-solubilising microorganisms often remains inconsistent because of poor survival and colonisation in competitive soil environments influenced by native microbial communities and non-living factors.13,14 Some bacteria, known as endophytes, survive in plant tissues without causing disease and represent a promising strategy owing to their improved persistence, intimate association with host plants, and capacity to enhance growth through nutrient mobilisation, stress resilience, and pathogen suppression.15
Glycine max, the plant, is a globally important leguminous crop recognised for its high protein content (approximately 39%) and oil content (about 20%), playing an important role in food security and agricultural economies around the world. In India, soybeans are a key oilseed crop, contributing substantially to edible oil production and export earnings, with Maharashtra among the leading producing states.16,17 Sustainable enhancement of soybean productivity is essential to meet increasing demand while preserving the health of the soil.
Accordingly, the primary objective of this research effort was to isolate and screen phosphate-solubilising microorganisms associated with soybean plants, followed by their evaluation for their multifunctional role in stimulating plant development. The chosen isolates were examined for phosphate solubilisation efficiency, making indole-3-acetic acid, making siderophores, and nitrogenase and ACC deaminase activities. The impact of bacterial inoculation on soybean growth was further investigated in pot culture. Overall, the results provide valuable evidence supporting the potential application of multifunctional PSB as sustainable biofertilisers for soybean cultivation.
PLANT MATERIAL
Ten healthy, symptom-free soybean plants were randomly collected from five different sites within the Shirala region during October and November 2022. The study area is located at an elevation of about 594 m above sea level and experiences an average temperature of 24 °C, with annual precipitation of about 1,078.90 mm, distributed over nearly 66 rainy days. Plants were carefully uprooted with intact root systems, thoroughly washed to remove adhering soil, and before further processing, the samples were kept at 4 °C.
Medium
Pikovskaya’s agar medium (PVK) was prepared and used for isolation. The comparable liquid PVK medium comprised the same components, but excluded agar, which was employed for isolate enrichment. Katznelson’s and Bose agar medium (KB) was prepared with glucose (10 g L-1), K2HPO4 (5 g L-1), CaCl2 (10 g L-1), agar (25 g L-1), and soil extract (390 mL L-1). The final volume was adjusted to 1 L with distilled water, and the pH was maintained at 7.0. The Luria-Bertani (LB) medium was prepared and autoclaved at 121 °C for 15 minutes before being used to cultivate, maintain, and propagate bacterial isolates.
Isolation of endophytes from soybean plant samples
Plant material from collected soybean plants, including roots, stems, and leaves, was removed from the refrigerator and rinsed with tap water to remove contaminants. After thorough washing, sterile filter paper absorbed any remaining moisture. About 1 gram of the plant materials was placed onto a Petri dish and washed with 75% ethyl alcohol for 30 seconds. After rinsing seven times with sterile water, excess water was blotted with sterile filter paper. Sterile surgical scissors cut the material into 2 mm × 2 mm pieces after surface sterilisation. A small amount of sterile quartz sand was used to grind these pieces in a sterile mortar. The mixture was processed to achieve a uniform blend. A sterile diluent was used to dilute the homogenate to concentrations of 10-1, 10-2, and 10-3 in sequence. The appropriate dilutions were used to disperse 100 µl across the medium.15 As a control, the sterilised water from the final rinse of the plant material was used. After 24 hours of room temperature incubation, the control and sample plates were removed. Colonies on control plates were used to test the efficiency of surface disinfection. Colonies on sample plates were observed and their characteristics recorded. Unique colonies were chosen for further investigation depending on characteristics. These isolates were purified on fresh medium until each showed a uniform colony morphology. Strains were purified and isolated using the streak-separation approach based on colony phenotype until each had a uniform colony shape.
Primary screening of phosphate solubility of the isolates
Primary screening of bacteria capable of phosphate solubilization was carried out using the spot inoculation method on Katznelson’s and Bose agar, wherein insoluble calcium phosphate CaHPO4 was formed in situ by the reaction between K2HPO4 and CaCl2, serving as the source of insoluble phosphorus. To evaluate the consistency of phosphate solubilization, the isolated bacterial strains were subcultured for successive generations and reassessed for their solubilization ability. The Solubilization Index (SI), calculated as shown in Equation (1), was used to evaluate the bacteria’s effectiveness in phosphate solubilisation.18
Solubilization index (SI) = Diameter of Colony + Diameter of Halo zone / Diameter of colony
Quantitative estimation of phosphate solubility of the isolates
The isolates were cultivated in Pikovskaya’s liquid medium enriched with tricalcium phosphate [Ca3(PO4)2] as an insoluble phosphorus source to evaluate phosphate-solubilizing activity. Flasks were inoculated with the bacterial culture and incubated at ambient temperature at pH 7.0 for a period of seven days. After incubation, cultures were centrifuged at a speed of 10,000 rpm for ten minutes to isolate the supernatant, in which soluble Phosphorus was quantified using the molybdenum–antimony–ascorbic acid method.19 The released Phosphorus was expressed in milligrams per litre (mg L-1).
Determination of nitrogenase activity
The potential isolate’s biological nitrogen fixation was evaluated using the quantitative test for acetylene reduction.20,21 Nitrogenase, the key enzyme that reduces atmospheric nitrogen to ammonia, can also reduce acetylene (C2H2) to ethylene (C2H4), which is the basis for this assay. The actively growing bacterial culture (0.2 mL) was inoculated into sterile nutrient broth, which was incubated at the optimum temperature for 12 hours. Prior to resuspending in nitrogen-free liquid medium, the cells underwent two washes with sterile distilled water after being collected by centrifugation. 2.4 mL of pure acetylene gas and 3 mL of the bacterial suspension (adjusted to 108 × CFU mL-1) were injected into a sterile 25 mL serum vial. The vials were placed into incubators at 30 °C for 12 hours. A gas chromatograph was used to measure the amount of ethylene produced by injecting 1 mL of headspace gas and comparing the peak area to that of a 1:1000-diluted standard ethylene solution. Nitrogenase activity was measured as micromoles (µmol) of ethylene generated per hour and the experiment was carried out in triplicate.
Activity of ACC deaminase
The activity of the ACC enzyme was tested according to the standard protocol.22 The isolate was grown in nutrient broth to acquire enough biomass, thereafter extracted using centrifugation at 6,000 rpm for 5 minutes, and rinsed twice with 0.85% sodium chloride solution. The cells were then placed in 8 mL of nutrient broth containing 4 mM aminocyclopropane-1-carboxylate as the sole nitrogen source. The suspension was put in an incubator at 28 °C with shaking at 120 rpm for 24 hours to induce enzyme activity. Bacterial cells were then point-inoculated onto rhizobial minimal medium (RMM) plates.22 The only nitrogen supply for the experimental plates was 1-ACC, while the control plates were without a nitrogen source. Colony diameters on ACC-amended and control plates were compared to assess ACC deaminase activity.23
IAA Production
A colorimetric test was used to quantify the synthesis of Indole-3-acetic acid (IAA), following the method reported by Gordon and Weber.24 The bacterial strain was grown for three days at 28 °C in TY broth with or without 490 µg mL-1 of L-tryptophan. After the incubation period, the cultures were centrifuged at 13,000 × g for 10 minutes. Afterwards, Salkowski’s reagent (4.5 g FeClƒ L-1 in 10.8 M H2SO4) was mixed with 2 mL of the supernatant. The absorbance was measured at 600 nm following a 30 minute incubation in an ambient temperature. Using a standard IAA calibration curve, the concentration of IAA was determined.25
Production of siderophore
Production of siderophores was tested using Chrome Azurol S (CAS) agar, according to the method reported by Schwyn and Neilands.26 After spot-inoculating the isolate onto CAS agar, the plates underwent incubation at 28 °C for 24-48 hours. The bacterial colony was surrounded by an orange halo, which was indicative of siderophore synthesis. The ratio of the halo diameter (D) to the colony diameter (d) was used to estimate the effectiveness of siderophore generation.
Physiological and biochemical characterisation of the isolates
The biochemical and physiological features of phosphate-solubilizing bacteria were studied in accordance with methods detailed in the Common Bacterial System Identification Manual and Bergey’s Manual of Systematic Bacteriology.27 Gram staining, hydrolysis of glucose and lactose, methyl red, Voges-Proskauer, formation of hydrogen sulfide, liquefaction of gelatin, use of citrate and malonate, denitrification, oxidase, and catalase tests were among the studies.
16S rRNA gene sequencing and phylogenetic analysis
The gene encoding 16S rRNA was amplified in a polymerase chain reaction (PCR) using primers 27F and 1482R. The genomic DNA of the selected isolate was extracted using a bacterial genomic DNA extraction kit.28 Amplification using polymerase chain reaction was carried out under standard conditions, and the resulting products were bidirectionally sequenced. The NCBI BLAST program was utilised to analyse sequence similarities. Multiple sequence alignment was performed using MUSCLE 3.7, and inadequately aligned segments were removed using Gblocks 0.91b.29,30 The HKY85 substitution model was employed to conduct phylogenetic analysis using PhyML 3.0, and the phylogenetic tree was generated using TreeDyn.31
Assessment of individual inoculation-based plant growth promotion preparation of inoculum
Individual colonies were introduced into LB broth and gently shaken at 28 °C for 12 hours. This starter culture was then introduced at 1% volume to 200 mL of fresh LB broth and grown for another 48 hours. The cells were collected and rinsed with sterile distilled water after incubation and standardized at a final population of 3 × 108 CFU per mL.32
The three-factor orthogonal design was used for pot culture experiments from August to November 2023 with five replications per treatment. The bacterial strain, the dilution ratio, and the inoculation method were the independent variables in the experiment.33 Treatments included rhizosphere irrigation, foliar spray, and a combination of rhizosphere irrigation and foliar spray. Height of the plant and diameter of the stem were measured prior to inoculation and after 90 days of treatment, along with root, stem, and leaf parameters.
Isolation of endophytes from soybean plant samples
Ten healthy soybean plants were collected from five locations in the Shirala region during October–November 2022, characterised by favourable agroclimatic conditions for soybean growth and microbial diversity. Proper sampling, cleaning, and storage ensured minimal contamination and preservation of viable plant-associated microorganisms. The successful recovery of bacterial isolates confirms that healthy soybean plants are reliable sources of beneficial endophytic and rhizospheric bacteria and that the adopted methodology is suitable for isolating plant growth–promoting and phosphate-solubilising bacteria. The successful isolation of bacterial strains from healthy soybean plants indicates stable plant–microbe interactions supported by favourable agroclimatic conditions, with healthy plant tissues serving as important reservoirs of beneficial endophytic and rhizospheric microorganisms that enhance nutrient acquisition and plant growth.15,34 These findings confirm that soybean plants create an ecological niche for isolating phosphate-solubilising microorganisms that boost plant growth with potential applications in sustainable agriculture.12,35
Morphological characterisation of the isolates
Ten endophytic bacterial isolates from soybean (Glycine max) exhibited distinct colony morphological characteristics when grown on culture media. The isolates showed variation in colony size, ranging from small (RSN, S2, S5) to medium (SWK, ASN, S3, S4, S7) and large colonies (S1, S6), indicating differences in growth rate and metabolic activity. Most isolates formed circular colonies, whereas a few, such as ASN, S3, and S6, displayed irregular shapes, suggesting morphological diversity within the bacterial population. Colony colour varied from white and creamy white to pale yellow and light cream, reflecting possible differences in pigment production among isolates. The majority of isolates exhibited entire margins; however, undulate and lobate margins were observed in ASN, S3, and S6, which further supports phenotypic diversity. Elevation patterns also differed: most isolates formed convex or raised colonies, whereas a few, such as RSN, S2, and S5, formed flat colonies.
Texture analysis revealed that most isolates were smooth. In contrast, isolates ASN and S3 showed rough or slightly rough textures, and S1 and S6 exhibited mucoid colonies, which may be associated with exopolysaccharide production and enhanced plant–microbe interactions. Overall, the observed variation in colony morphology among the ten isolates indicates the presence of diverse endophytic bacterial communities within soybean plants. These morphological differences provided a basis for preliminary differentiation and selection of isolates for further physiological, biochemical, and molecular characterisation, particularly to identify strains with phosphate-solubilising and plant growth–promoting potential.
The observed diversity in colony morphology among the ten endophytic bacterial isolates reflects substantial phenotypic and metabolic variability within soybean-associated microbial communities. Variations in colony size, shape, colour, margin, elevation, and texture are widely recognised as indicators of functional diversity and adaptive potential among plant-associated bacteria.15,35 The presence of mucoid colonies in some isolates suggests exopolysaccharide production, which facilitates root colonisation, biofilm formation, and enhanced plant–microbe interactions.12,36 Such morphological heterogeneity provides an important basis for preliminary differentiation of isolates before detailed physiological, biochemical, and molecular characterisation. Similar approaches have been successfully applied to identify efficient phosphate-solubilising and plant growth–promoting bacteria from soybean and other crop plants.10,34 This study found morphological diversity, suggesting functionally diverse endophytic bacteria for sustainable agriculture.
Screening of phosphate solubility of the isolates
Primary screening of phosphate-solubilising bacteria on Katznelson’s and Bose agar media revealed clear halo zones around all 10 isolates, indicating their ability to solubilise insoluble phosphate (Figure 1). The colony diameter, halo zone diameter, and Solubilization Index varied greatly. Among the isolates, SWK showed the highest halo zone diameter (12.5 mm) and the maximum SI value (3.50), demonstrating superior phosphate-solubilising efficiency compared to the other isolates. RSN and ASN exhibited moderate phosphate solubilisation with SI values of 2.78 and 2.63, respectively, while isolates S4, S2, and S7 also showed appreciable activity with SI values ranging from 2.35-2.50. In contrast, isolates S1, S3, S5, and S6 recorded comparatively lower SI values (2.17 to 2.27), suggesting weaker phosphate-solubilising potential.
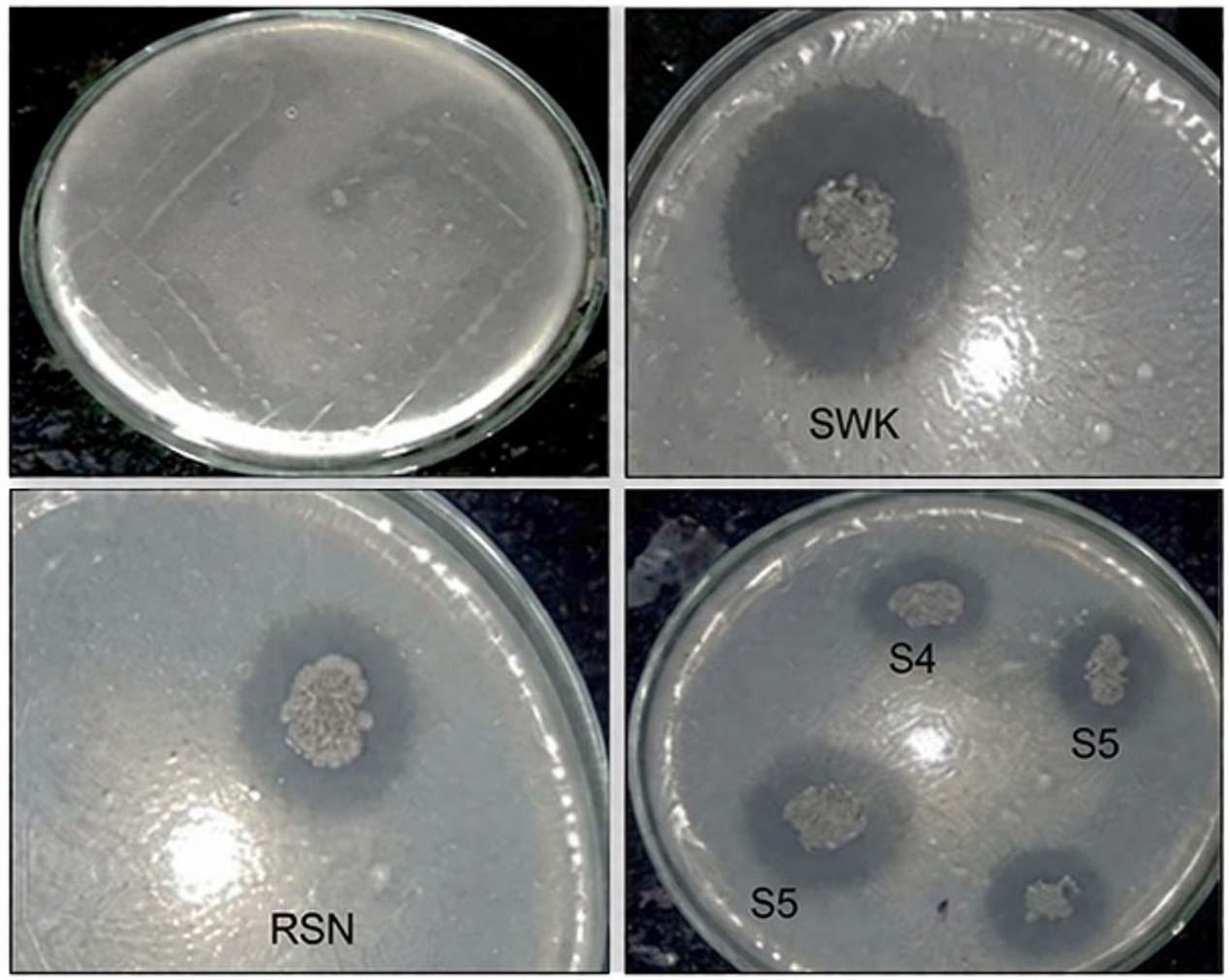
Figure 1. Primary screening of phosphate-solubility of the isolates on Katznelson’s and Bose agar media showing halo zone formation
The observed differences in SI values reflect variability in isolates’ ability to release organic acids or other metabolites that solubilise insoluble phosphate. The consistently higher solubilisation efficiency and stable halo formation of isolate SWK during repeated sub-culturing highlighted its reliability and effectiveness, justifying its selection for further physiological, biochemical, and molecular characterisation. Overall, the results confirm that primary screening is essential for identifying efficient phosphate-solubilising bacterial strains with potential biofertiliser applications.
The SWK isolate inoculation led to a considerable increase in soluble Phosphorus (704.7%) compared to the control (P < 0.05) clearly demonstrating the isolate’s strong phosphate-solubilising ability under liquid culture conditions. The formation of halo zones by all isolates confirms their phosphate-solubilising ability. In contrast, variations in halo diameter and Solubilization Index (SI) reflect strain-specific differences in phosphate mobilisation mechanisms like organic acid generation and proton extrusion.10 Among the isolates, SWK exhibited the highest and most stable SI value and showed a significant increase in soluble Phosphorus under liquid culture conditions, indicating superior phosphate-solubilising efficiency.15,34 These results highlight the importance of combined qualitative and quantitative screening for identifying efficient PSB and support the selection of SWK as a promising biofertiliser candidate for sustainable agriculture.12,33
Determination of nitrogenase activity
The selected isolate (SWK)’s nitrogenase activity was tested using the acetylene reduction assay after 12 hours of incubation in nitrogen-free media. The SWK isolate showed distinct nitrogenase activity, as indicated by the production of ethylene (4.86 ± 0.32 µmol C2H4 per hour), whereas no ethylene production was detected in the uninoculated control. The absence of ethylene in the control confirms that the observed activity was due to the bacterial isolate’s metabolic activity. The consistent ethylene production across replicates indicates stable, reliable nitrogenase activity in the SWK isolate. This ability to fix atmospheric nitrogen suggests that the isolate can improve nitrogen availability for plants, thereby enhancing plant growth. When combined with its phosphate-solubilising potential, the nitrogen-fixing capability of the SWK isolate highlights its multifunctional role as a plant growth–promoting bacterium. Such traits are particularly beneficial for soybean cultivation and support the potential application of the SWK isolate as an effective biofertiliser for sustainable agriculture.
The detection of ethylene production by the SWK isolate in the acetylene reduction assay confirms its active nitrogenase enzyme system and ability to fix atmospheric nitrogen, a fundamental characteristic of plant growth-promoting microorganisms. A lot of research has shown that free-living and associative microorganisms that fix nitrogen can make nitrogen more available and improve crop growth, particularly in leguminous plants such as soybean.37,38 The lack of ethylene synthesis in the uninoculated control further confirms that the observed activity was entirely related to bacterial metabolism. One crucial feature for the SWK isolate’s possible use in the field is its metabolic stability, which is shown by its consistent nitrogenase activity across replicates. When combined with its phosphate-solubilising potential, the nitrogen-fixing capability of the SWK isolate highlights its multifunctional role as a plant growth–promoting bacterium. Such traits are particularly beneficial for soybean cultivation and support the potential application of the SWK isolate as an effective biofertiliser for sustainable agriculture. 12,34
Activity of ACC deaminase
A comparison was conducted on the bacterial growth of the SWK isolate on rhizobial minimal medium, both with and without ACC as the single nitrogen source, to evaluate its 1-aminocyclopropane-1-carboxylate deaminase activity. After three days of incubation on ACC-supplemented media, the SWK isolate grew significantly, with a colony diameter of 4.8 ± 0.4 mm. In contrast, minimal growth was observed on control plates lacking a nitrogen source (1.2 ± 0.3 mm). The SWK isolate uses ACC as a source of nitrogen, as seen by the drastic increase in colony diameter on ACC-containing plates suggesting deaminase (ACC) activity. Degrading ACC, the immediate precursor of ethylene, via such specific deaminase reduces stress-induced ethylene inhibition in plants. The SWK isolate’s positive deaminase (ACC) activity may boost plant development, especially under stress.
The SWK isolate’s capacity to grow on ACC alone proves its ACC deaminase activity, a critical characteristic of stress-alleviating plant growth-increasing bacteria. ACC deaminase degrades 1-aminocyclopropane-1-carboxylate to lower plant ethylene levels and mitigate ethylene-mediated growth inhibition under biotic and abiotic stress.39,40 The significantly greater colony growth of SWK on ACC-supplemented medium compared to the nitrogen-free control further validates its functional ACC utilisation capacity. The presence of ACC deaminase activity, together with phosphate solubilisation and nitrogen fixation, highlights the multifunctional plant growth–promoting potential of the SWK isolate. Such multifunctionality is a critical criterion for selecting effective bioinoculants that enhance plant growth and stress tolerance in sustainable agricultural systems.12,34
IAA Production
A colorimetric assay in TY broth, both with and without L-tryptophan, evaluated the indole-3-acetic acid (IAA) biosynthesis of the SWK isolate. No formation of IAA was found in the control group. The SWK isolate produced a measurable amount of IAA (6.4 ± 0.5 µg/mL) even in the absence of L-tryptophan, indicating its inherent ability to synthesise IAA. However, supplementation with L-tryptophan (490 µg/mL) significantly enhanced IAA production to 28.7 ± 1.8 µg/mL, demonstrating a tryptophan-dependent IAA biosynthetic pathway. L-tryptophan increases IAA production, suggesting the SWK isolate may efficiently use tryptophan as an auxin precursor. IAA is known to stimulate root elongation, lateral root formation, and overall plant growth. Thus, the SWK isolate’s ability to create large levels of IAA, especially under tryptophan-supplemented conditions, supports its potential as a multifunctional plant growth–promoting bacterium. The SWK isolate’s capacity to generate indole-3-acetic acid (IAA) both with and without L-tryptophan suggests the presence of numerous IAA biosynthetic pathways, which is a frequent trait of efficient plant growth-promoting bacteria. The significantly enhanced IAA production under tryptophan-supplemented conditions confirms the involvement of a tryptophan-dependent auxin biosynthesis pathway, as reported in rhizosphere and endophytic bacteria.35,41 Such tryptophan-mediated IAA synthesis is known to be influenced by root exudates, allowing bacteria to respond dynamically to plant signals. Plant-associated bacteria create IAA, which stimulates root elongation, lateral root creation, and root hair development, enhancing plant nutrition and water intake.12,34 The substantial IAA production observed in the SWK isolate, together with its phosphate-solubilising, nitrogen-fixing, and ACC deaminase activities, highlights its multifunctional plant growth–promoting potential. These combined traits support SWK’s suitability as an effective biofertiliser candidate for sustainable agricultural practices.
Production of siderophore
Chrome Azurol S (CAS) agar showed an orange halo around bacterial colonies, indicating siderophore synthesis by the identified isolate. The SWK isolate produced a distinct orange halo with a diameter of 9.8 ± 0.6 mm around colonies measuring 4.5 ± 0.4 mm, yielding a D/d ratio of 2.18. The control showed no halo formation, indicating no siderophore production.
The SWK isolate’s distinct orange halo and high D/d ratio suggest robust iron-chelation. Siderophore production is an important plant growth–increasing trait, as it enhances iron availability to plants and suppresses the growth of phytopathogens by limiting their access to iron. The SWK isolate forms an orange halo on CAS agar, confirming its impressive siderophore-producing and iron-chelating abilities. Siderophore production enhances iron availability to plants and suppresses phytopathogens by limiting their access to iron, thereby improving plant growth and health.34,42 Siderophore activity of the SWK isolate, in addition to the synthesis of IAA, the deaminase ACC activity, nitrogen fixation, and phosphate solubilization, is a potential bioinoculant for sustainable agriculture due to its multifunctionality, which promotes the development of plants.12,15
Physiological and biochemical characterisation of the isolate
Physiological and biochemical characterisation of the phosphate-solubilising bacterial isolate SWK revealed distinct metabolic and enzymatic traits. The isolate was characterized as a Gram-negative, shaped like rod bacterium and showed positive reactions for glucose and lactose hydrolysis, indicating its ability to utilise carbohydrates efficiently. Positive Voges Proskauer, citrate, malonate, gelatin liquefaction, oxidase, catalase, and denitrification tests further reflect its metabolic versatility and adaptability to diverse soil environments.10,34 The negative methyl red and hydrogen sulfide tests helped differentiate the isolate from other closely related bacteria. Overall, the biochemical profile of SWK is characteristic of efficient plant growth–promoting and phosphate-solubilising bacteria and supports its identification and further confirmation by molecular analysis.12,15
16S rRNA gene sequencing and phylogenetic analysis
The molecular characterization was performed through 16S rRNA gene sequencing to achieve accurate taxonomic identification. The 16S rRNA gene sequence obtained from isolate SWK was deposited to the DNA Data Bank of Japan (DDBJ) under the designation Enterobacter hormaechei strain SWK, with accession number LC923381. The phylogenetic tree based on 16S rRNA gene sequences shows that isolate SWK clusters within the Enterobacter hormaechei group (Figure 2). The phylogenetic analysis strongly supports that the isolate SWK belongs to Enterobacter hormaechei and is closely related to other members of the E. hormaechei lineage within the Enterobacter cloacae complex.
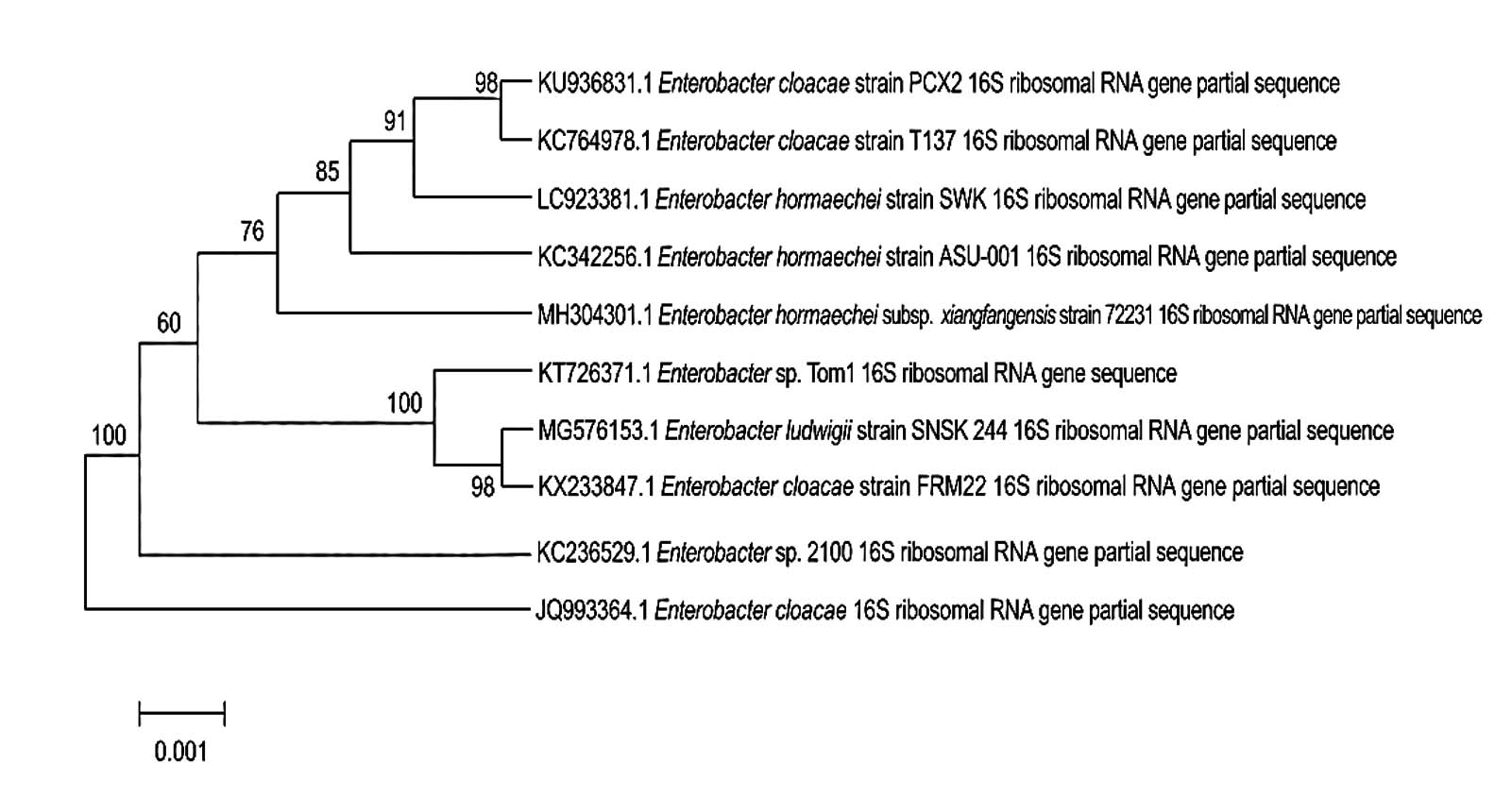
Figure 2. Phylogenetic relationship of the SWK isolate with closely related Enterobacter species based on nucleotide sequence analysis
Assessment of individual inoculation-based plant growth promotion preparation of inoculum
In pot culture experiments, inoculation with the bacterial isolates considerably affected soybean (Glycine max) height, stem diameter, root length, and biomass. (Figures 3 and 4). Growth responses varied by bacterial isolate, dilution ratio, and inoculation method.
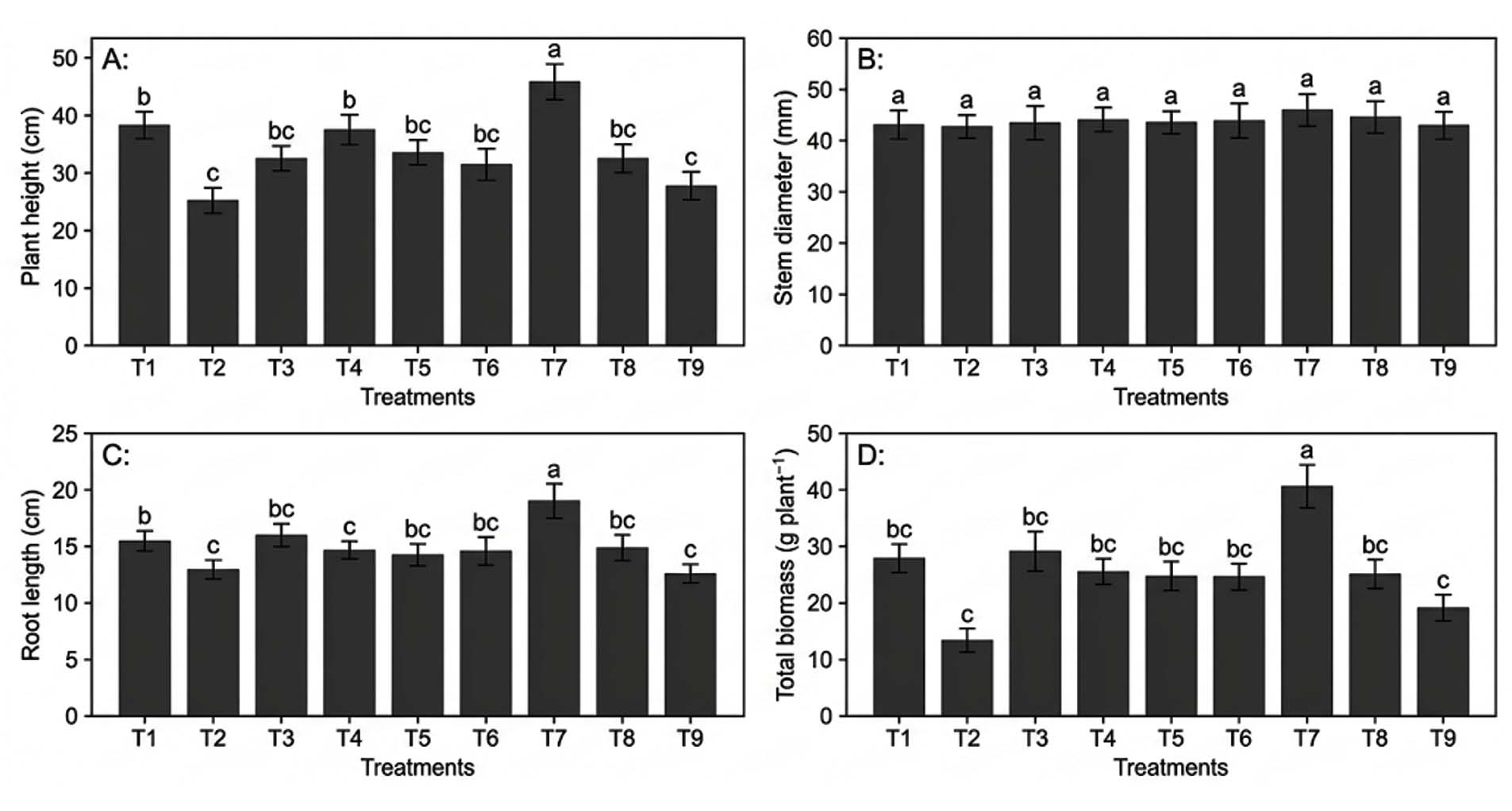
Figure 3. The impact of the bacterial inoculation treatments on soybean (Glycine max) growth metrics in pot culture A) Plant height (cm), B) Stem diameter (mm), C) Root length (cm), and D) Total Biomass g/plants. (All values are the means of three replicates ± SD. Different small case letters statistically significant differences.)

Figure 4. Effect of bacterial inoculation treatments on growth attributes of soybean (Glycine max) under pot culture conditions
Plant height ranged from 34.9 ± 1.6 cm to 46.8 ± 2.2 cm. Treatments with the SWK isolate consistently produced taller plants than those with RSN or ASN, indicating isolate-specific differences in growth-promoting efficiency. The superior performance of the SWK isolate is likely due to its multifunctional traits, including phosphate solubilisation and phytohormone production.
Stem diameter showed a similar trend, with the highest value recorded in T7 (7.4 ± 0.4 mm) and the lowest in T2 (5.1 ± 0.2 mm). Increased stem thickness under SWK treatments reflects improved structural development and nutrient translocation. Root length was also maximised under T7 (19.8 ± 1.3 cm), suggesting enhanced root elongation and branching, possibly mediated by bacterial indole-3-acetic acid production.
Total biomass accumulation varied significantly among treatments, with T7 producing the highest biomass (13.8 ± 0.7 g plant-1), followed by T1 and T4, while the lowest biomass was observed in T9 (9.1 ± 0.4 g plant-1). Biomass improvement integrates the cumulative effects of enhanced nutrient availability, stress mitigation, and improved physiological performance.
Inoculation treatments had a highly significant effect on plant height, stem diameter, root length, and total biomass of soybean (Glycine max), as determined by one-way analysis of variance (ANOVA) (P < 0.001). This confirms that bacterial isolate, dilution ratio, and inoculation method significantly influenced plant growth responses.
Across all parameters, treatments with SWK consistently outperformed those with RSN and ASN, highlighting isolate-specific differences in plant growth–promoting efficiency. Moreover, the combined rhizosphere irrigation and foliar spray (RF) method resulted in significantly greater growth enhancement than individual application methods, emphasising the importance of effective root–microbe interactions. The low standard deviation values and clear separation of treatment mean further validate the robustness of the experiment and the suitability of the data for ANOVA and post hoc comparisons.
Soybean plants inoculated with the SWK isolate grew taller, had wider stems, and produced more biomass than their uninoculated counterparts. Many studies have shown that phosphate-solubilizing and plant growth-promoting bacteria boost growth. and are mainly attributed to improved nutrient availability and phytohormone-mediated root development.10,34 The superior performance of the combined rhizosphere irrigation and foliar spray (RF) treatment suggests that simultaneous root and leaf colonisation enhances bacterial establishment and nutrient uptake efficiency, thereby improving plant physiological responses.12,43 The isolate SWK appears to be a promising plant growth–promoting bacterium for soybean growing in pot culture.
The present study successfully isolated and characterised endophytic and rhizospheric bacteria from healthy soybean plants, identifying the isolate SWK as a highly efficient and multifunctional plant growth–promoting bacterium. The isolate SWK exhibited strong phosphate-solubilising ability both qualitatively and quantitatively, along with stable nitrogenase activity, ability to degrade ACC, synthesis of indole-3-acetic acid, and siderophore synthesis, indicating its potential to enhance nutrient availability, reduce stress-induced ethylene effects, and promote overall plant growth. Experimental evidence from the fields of physiology, biochemistry, and molecular biology points to the species Enterobacter hormaechei, which belongs to the family Enterobacter cloacae.
The pot culture experiments further demonstrated that inoculation with SWK significantly improved soybean growth factors such as plant height, diameter of stems, and amount of biomass, with the combined rhizosphere irrigation and foliar spray method showing the most pronounced effect. Collectively, these findings highlight the effectiveness and robustness of Enterobacter hormaechei strain SWK as a promising biofertiliser candidate. The multifunctional characteristics of SWK enhance its use in sustainable soybean cultivation and establish a robust basis for forthcoming field evaluations and formulation advancements.
ACKNOWLEDGMENTS
This study was made possible with the support and facilities provided by the Departments of Microbiology at VNBN College and DABN College.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
Both authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Food and Agriculture Organization of the United Nations. The State of Agricultural Commodity Markets 2022. The Geography of Food and Agricultural Trade: Policy Approaches for Sustainable Development; 2022.
Crossref - Smil V. Phosphorus in the environment: natural flows and human interferences. Annu Rev Energy Environ. 2000;25(1):53-88.
Crossref - Cordell D, Drangert JO, White S. The story of phosphorus: global food security and food for thought. Glob Environ Change. 2009;19(2):292-305.
Crossref - Elser JJ, Bennett E. A broken biogeochemical cycle. Nature. 2011;478(7367):29-31.
Crossref - Vance CP, Uhde-Stone C, Allan DL. Phosphorus acquisition and use. New Phytol. 2003;157(3):423-447.
Crossref - Lynch JP. Root Phenes for Enhanced Soil Exploration and Phosphorus Acquisition: Tools for Future Crops. Plant Physiol. 2011;156(3):1041-1049.
Crossref - Hinsinger P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil. 2001;237(2):173-195.
Crossref - Rodrı́Guez H, Fraga R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol Adv. 1999;17(4-5):319-339.
Crossref - Goldstein AH. Recent Progress in Understanding the Molecular Genetics and Biochemistry of Calcium Phosphate Solubilization by Gram Negative Bacteria. Biol Agric Hortic. 1995;12(2):185-193.
Crossref - Glick BR. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica (Cairo). 2012;2012:963401.
Crossref - Sharma SB, Sayyed RZ, Trivedi MH, Gobi TA. Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus. 2013;2(1):587.
Crossref - Backer R, Rokem JS, Ilangumaran G, et al. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front Plant Sci. 2018;9:1473.
Crossref - Bashan Y, de-Bashan LE, Prabhu SR, Hernandez JP. Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998–2013). Plant Soil. 2014;378(1-2):1-33.
Crossref - Trivedi P, Leach JE, Tringe SG, Sa T, Singh BK. Plant–microbiome interactions: from community assembly to plant health. Nat Rev Microbiol. 2020;18(11):607-621.
Crossref - Hardoim PR, van Overbeek LS, Berg G, et al. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol Mol Biol Rev. 2015;79(3):293-320.
Crossref - Directorate of Economics and Statistics. Agricultural statistics at a glance. Ministry of Agriculture & Farmers Welfare. 2021. https://agriwelfare.gov.in/Documents/CWWGDATA/Agricultural_Statistics_at_a_Glance_2021_1.pdf
- Prashnani M, Dupare B, Vadrevu KP, Justice C. Towards food security: exploring the spatio-temporal dynamics of soybean in India. PLOS ONE. 2024;19(5):e0292005.
Crossref - Nautiyal CS. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol Lett. 1999;170(1):265-270.
Crossref - Murphy J, Riley JP. A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta. 1962;27:31-36.
Crossref - Hardy RWF, Holsten RD, Jackson EK, Burns RC. The Acetylene-Ethylene Assay for N2 Fixation: Laboratory and Field Evaluation. Plant Physiol. 1968;43(8):1185-1207.
Crossref - Dobereiner J, Marriel IE, Nery M. Ecological distribution of Spirillum lipoferum Beijerinck. Can J Microbiol. 1976;22(10):1464-1473.
Crossref - Glick BR. The enhancement of plant growth by free-living bacteria. Can J Microbiol. 1995;41(2):109-117.
Crossref - Glick BR, Cheng Z, Czarny J, Duan J. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur J Plant Pathol. 2007;119(3):329-339.
Crossref - Gordon SA, Weber RP. Colorimetric Estimation of indoleacetic Acid. Plant Physiol. 1951;26(1):192-195.
Crossref - Patten CL, Glick BR. Role of Pseudomonas putida Indoleacetic Acid in Development of the Host Plant Root System. Appl Environ Microbiol. 2002;68(8):3795-3801.
Crossref - Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987;160(1):47-56.
Crossref - Bergey DH, Whitman WB, Goodfellow M, et al., ed. Bergey’s Manual of Systematic Bacteriology. 2nd ed. Springer New York, NY; 2012.
Crossref - Weisburg WG, Barns SM, Pelletier DA, Lane DJ. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol. 1991;173(2):697-703.
Crossref - Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792-1797.
Crossref - Talavera G, Castresana J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst Biol. 2007;56(4):564-577.
Crossref - Dereeper A, Guignon V, Blanc G, et al. Phylogeny.fr. Nucleic Acids Res. 2008;36(Web Server Issue):W465-W469.
Crossref - Somasegaran P, Hoben HJ. Handbook for Rhizobia. Springer New York, NY;1994.
Crossref - Vessey JK. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 2003;255(2):571-586.
Crossref - Berg G, Rybakova D, Grube M, Köberl M. The plant microbiome explored: implications for experimental botany. J Exp Bot. 2015;67(4):995-1002.
Crossref - Santoyo G, Moreno-Hagelsieb G, Orozco-Mosqueda MDC, Glick BR. Biofilm formation in plant–microbe associationss. Microbiol Res. 2016;183:92-99.
Crossref - Ramey BE, Koutsoudis M, von Bodman SB, Fuqua C. Biofilms. Curr Opin Microbiol. 2004;7(6):602-609.
Crossref - Peoples MB, Herridge DF, Ladha JK. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? Plant Soil. 1995;174(1-2):3-28.
Crossref - Zahran HH. Rhizobium-Legume Symbiosis and Nitrogen Fixation under Severe Conditions and in an Arid Climate. Microbiol Mol Biol Rev. 1999;63(4):968-989.
Crossref - Glick BR, Penrose DM, Li J. A Model For the Lowering of Plant Ethylene Concentrations by Plant Growth-promoting Bacteria. J Theor Biol. 1998;190(1):63-68.
Crossref - Penrose DM, Glick BR. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria Physiol Plant. 2003;118(1):10-15.
Crossref - Spaepen S, Vanderleyden J, Remans R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol Rev. 2007;31(4):425-448.
Crossref - Neilands JB. Siderophores: Structure and function of microbial iron transport compound. J Biol Chem. 1995;270(45):26723-26726
- Hoffmann H, Stindl S, Ludwig W, et al. Reassignment of Enterobacter dissolvens to Enterobacter cloacae as E. cloacae subspecies dissolvens comb. nov. and emended description of Enterobacter asburiae and Enterobacter kobei. Syst Appl Microbiol. 2005;28(3):196-205.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.