


ISSN: 0973-7510
E-ISSN: 2581-690X
Endophytes are a group of bacteria live in plant tissues, and have been numerously reported to promote plant growth. The aim of this study was to observe bacterial endophytes from blackboard tree (Alstonia scholaris) in inhibiting pathogenic fungal growth on chili plants, promoting plant growth and producing indole acetic acid, as well as in solubilizing phosphate. Sixteen endophytic bacteria were isolated from blackboard tree root and leaf samples, in which two, A1 and A3, out of sixteen showed to highly inhibit pathogenic fungi Fusarium sp. and Colletrotichum sp., to produce indol acetic acid, and to solubilize phosphate. 16S rRNA gene sequence showed that A1 was relativey closed to Carnobacterium gallinarum strain DSM4847 and A3was relatively closed to Carnobacterium maltaromaticum strain JCM 1154 with similarity of 84 and 78%, respectively.
Alstonia scholaris, blackboard tree, chili, endophytes, indole acetic acid, phosphate
Modern agriculture are heavily dependent on chemical compounds such as fertilizers and pesticides that may cause environmental stress, reducing soil fertility, and even causing environmental instability due to the change in energy flow in the ecosystem (Amilia et al., 2016). Pesticides residue on agricultural products can also decrease product quality that causes the selling price of these products to decrease (Supardi, 1994). Thus, the urge for precise, cheap, safe, and innovative control is now in demand. To address this issue, utilization of endophytic microorganisms producing phytohormones such as indole acetic acid (IAA) that can stimulate plant growth, phosphate solubilizing microorganisms, as well as the use of microorganisms to control plant pests and diseases. In general, the role of microbes is to increase the availability of nutrients in the soil, decomposition and mineralization of organic matter in the soil, rhizosphere-endophytic bacteria promoting plant growth by forming rhizosphere – endophytes symbiosis and protect plant roots from pathogenic microbes, and plays a role as a natural pest and disease control agents. Various chemical reactions in the soil also occur with the help of soil microbes.
Microbes enter plant tissue in various ways, such as through injuries to plant tissue, leaf stomata, or through the pores of the roots. The ability of bacteria to penetrate the plant’s internal tissue can be due to the extracellular enzyme produced by endophytic bacteria (Saraswati et al., 2004). Endophytic bacteria can produce bioactive compounds whose character is similar to the compounds produced by their host plants (Eliza et al., 2007). A number of endophytic microbes are known to significantly produce phytohormones, especially IAA (Tan and Zou, 2001). Host plants infected with endophytes have many advantages, such as accelerated growth and increased resistance to drought and pest attacks. The role of host plants is to provide the nutritional needs for endophytic microbes. The condition of host plants, soil conditions i.e. temperature and humidity can greatly affect the number and type of endophytic microbes. The presence of endophytic microbes in plant tissue can also help plants to compete in the nature.
Blackboard tree (Alstonia sp) is categorized as a fast growing plant native to Indonesia (Pamungkas et al., 2006). The genera Alstonia itself belongs to the family Apocynaceae, similar to Rauvolfia canescens, Rauvolfia serpentine, Alstonia boonei, and Vinca rosea, which had been commercially produced as herbal medicine in China and India (Pratyush et al., 2011). A study on endophytic bacteria isolated from blackboard tree leaf (Alstonia scolaris) has previously been conducted. This study showed that the endophytic bacteria had a potential to produce antibacterial compounds against Escherichia coli and Staphylococcus aureus (Fatiqin, 2009). However, no study was carried out in their ability to produce indol acetic acid (IAA), to solubilize phosphate, and to inhibit of pathogenic fungi in chili.
Isolation of Endophytic Bacteria from Blackboard Tree
Endophytic bacterial isolation from roots and leaves of blackboard tree was performed as described by Radu et al. (2002). Samples were immediately washed with water to remove dirts, then dried and wrapped in newspaper and put into a plastic bag containing ice blocks (Zinniel et al., 2002). Samples were then brought to the laboratory for isolation. Samples were initially washed with running water. Sterilization of the root surface area was done by immersing the following solution successively: 75% ethanol for 5 minutes, 5.3% sodium hypochlorite solution for 5 minutes, and 75% ethanol for 30 seconds. Subsequently, the roots of the plant were rinsed with sterile distilled water then dried with sterile filter paper. Afterwards, the root part is cut to 1-1.5 cm, and each piece of root was placed on NA medium mixed with antifungal ketoconazole (0.3 gram/100 ml) incubated for ± 5 days. Similar procedure was done for leaf samples. The colonies were subcultured into a new NA medium to be purified.
Characterization of Morphology and Biochemistry of Bacterial Endophytes
Bacterial isolates were morphologically characterized by observing their colony shape, color, elevation and edge. Gram staining was performed to observe their cell shape, arrangement and bacterial gram reaction. Biochemical tests including starch hydrolysis, gelatin hydrolysis, citrate utilization test, hydrogen sulfide test, and motility test were conducted to observe their biochemical activity.
Antagonistic Assay of Endophytic Bacteria against Pathogenic Fungi
Inhibition of endophytic bacterial against pathogenic fungi was carried out in vitro. Pathogenic fungi used in this study were Colletotrichum sp. and Fusarium sp. Active growing mycellia of the pathogenic fungus was taken using cork borer, and subsequently inoculated in the middle of Mueller’s Hinton Agar medium. Endophytic bacteria were carefully inoculated using a sterile toothpick 3 cm away from the pathogenic fungal cultures, Cultures were incubated for 3 days.
Growth of Endophytic Bacterial Isolates
Endophytic bacteria cultures previously grown on nutrient agar (NA) were inoculated into5 ml of sterile physiological saline solution. Bacterial suspension of 5% was introduced into 50 ml of sterile Luria Bertani (LB) containing tryptophan, and spectrophotometry measurement was done at 600 nm wavelength. The initial absorbance of LB + tryptophan was used as blank, and initial measurement of LB containing bacteria was conducted aseptically. Culture was incubated on orbital shaker at 27oC of 100 rpm. The absorbance of the culture was measured every 4 hours to know the bacterial growth until it reached its stationary phase.
Solubilizing Phosphate Assay of Endophytic Bacteria
Endophytic bacterial isolates were assayed for their ability in solubilizing phosphate. This was done by inoculating the bacterial isolate on Pikovskaya agar aseptically using sterile toothpick. Culture was incubated for 7 days. Phosphate Solubilization Index (PSI) was determined by comparing the diameter of clear zone formed in the agar against the diameter of the growing bacterial colony.
Phosphate Solubilization Index (PSI) = diameter of clear zone/diameter of colony
Assay of Endophytic Bacterial Isolatesin Producing IAA
An assay on endophytic bacterial isolate ability to produce IAA was carried out as described by Dewi et al. (2016). Bacterial isolates were subcultured on NA for 24 hours. A5 ml of culture suspension was adjusted to 0.5 Mc Farland standard (H”108cell/ml). Suspension of 5%was inoculated into LB broth + tryptophan and then incubated in orbital shaker at 27oC of 100 rpm. Measurement of IAA was done every 4 hours using centrifugation method at 5000 rpm for 10 minutes. Analysis of IAA production was determined using spectrofotometric method. A 0.5 ml of Salkowsky reagent was dropped into 2 ml of supernatant atratio of 4:1. Solution was incubated at ambient temperature for 15 minutes. Its absorbance was measured using spectrophotometer at 530 nm. Measurement of IAA curve was terminated after the bacterial culture reached stationary phase.
Application of Potential Endophytic Bacteria on Red Chilli Seeds
Suspension of potential endophytic bacteria with cell density of 108 CFU/ml was inoculated to LB broth to make 5% culture suspension. Culture was incubated at orbital shaker for 4 hours at ambient temperature. Chili root were soaked into the suspension for 15 minutes and planted in a 3 kg-polybag previously filled with soil compost. Observation was done 30 days after plantation by measuring plant height, number of leaves, fresh weight, dry weight, and number of chlorophylls (Dey et al., 2004).
Identification of Endophytic Bacteria Based on 16S rRNA
Extraction of DNA of potential endophytic bacteria was done using freeze and thaw method. A 1.5 ml of pure bacterial culturewas centrifuged for 10 minutes. Supernatant was removed. Cell pelletwas added aseptically with 100 ¼L of steriled water and suspended. Cell suspension was subsequently frozen at -10° C and then heated at 90°C for 10 minutes for 5-10 times (Nusyirwani, 2007).
To make a 25 µL of PCR reaction solution, the following materials were added into 0.2 mL eppendorf tube: 12,5 µL of Master Mix 2X GoTaqGreen (Promega), 1 µL of forward primer (10 pmol), 1 µL of reverse primer 10 (pmol), 2 µL of template DNA, and 8,5 µL of nuclease free water. PCR machine was programmed and run using the following temperature: pradenaturation at 94oC for 2 minutes, denaturation at 92oC for 30 seconds, annealing at 55oC for 30 seconds, primer elongation at 72oC for 1 minute, and post–PCR at 72oC for 5 minutes. This process was run for 30 cycles. Amplification of 16S rRNA gene of broken cell was conducted using Polymerse Chain Reaction (PCR) machine (Labcycler Gradient) with a 16S rRNA-specific universal primer for prokaryotes, 63f (5′- CAG GCC TAA CAC ATG CAA GTC – 3 ‘) and 1387r (5’ – GGG CGG WGT GTA CAA GGC 3 ‘)(Marchesi et al., 1998). PCR product was analyzed with electrophoresis. Analysis was performed at 80 volts and 400 mA for 90 minutes, and then visualized using UV-transluminator.
The amplified DNA was commerciallypurified and sequenced. DNA sequence was compared to that of GenBank database of The National Center for Biotechnology Information (NCBI), using Basic Local Alignment Search Tool (BLAST).
Characterization of Endophytic Bacteria from Roots and Leaves of Blackboard Plant
Sixteen endophytic bacteria were isolated in this study, of which 8 isolates were obtained from the root and 8 isolates were from the leaves of blackboard tree. Other isolated 9 isolates of endophytic bacteria from blackboard tree leaves (Fatiqin, 2009). Characterization on the bacterial isolates was carried out, including cell morphology, bacterial colonies and biochemical tests (Table 1). Of this characterization, it showed that all isolates might be different.
Table (1):
Morphological and Biochemical Characteristics of Endophytic Bacterial Isolates of Blackboard Tree
| Source | Isolate code | Colony morphology | Gram staining | Biochemical Traits | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Shape | Edge | Elevation | Color | Starch metabolism | Gelatin hydrolysis | Citrate metabolism | H2S production | Motility | |||
| Root | A1 | Circular | Entire | Flat | Cream | Positive | – | – | – | + | + |
| A2 | Circular | Entire | Flat | Cream | Negative | + | – | + | + | + | |
| A3 | Circular | Entire | Raised | Cream | Positive | – | – | + | + | + | |
| A4 | Irregular | Undulate | Flat | Pink | Negative | – | – | + | + | + | |
| A5 | Irregular | Entire | Convex | Cream | Negative | – | – | + | + | + | |
| A6 | Circular | Entire | Flat | Cream | Negative | – | – | + | + | + | |
| A7 | Irregular | Undulate | Flat | Pink | Negative | + | – | + | + | + | |
| A8 | Circular | Entire | Flat | Cream | Positive | – | – | – | – | + | |
| Leaves | D1 | Irregular | Undulate | Flat | Cream | Negative | – | – | + | + | + |
| D2 | Circular | Entire | Convex | Cream | Positive | – | – | + | + | + | |
| D3 | Circular | Entire | Convex | Cream | Negative | + | – | + | + | + | |
| D4 | Circular | Entire | Convex | Cream | Negative | – | + | + | + | + | |
| D5 | Circular | Entire | Flat | Cream | Negative | – | + | + | + | + | |
| D6 | Circular | Entire | Convex | Cream | Negative | – | + | + | + | + | |
| D7 | Circular | Entire | Convex | Cream | Negative | – | + | + | + | + | |
| D8 | Irregular | Undulate | Flat | Cream | Negative | – | + | + | + | + | |
Assay on Endophytic Bacteria of Blackboard Tree to Inhibit Colletotrichum sp. and Fusarium sp. Growth
The ability of endophytic bacterial isolates to inhibit fungal growth is determined by the presence of inhibition zone at the intersection area between bacterial isolates and pathogenic fungi. The ability of endophytic bacteria to inhibit pathogenic fungi is based on their ability to produce metabolites such as cell wall degradation enzymes or other antifungal compounds (Ramamoorthy et al., 2002).
Two out of 16 isolates were observed to have the ability to inhibit both pathogenic fungi, ten were able to inhibit only either one of the pathogenic fungi, while the other four was not able to inhibit pathogenic fungi (Table 1). Previous study on endophytic bacteria from blackboard tree leaf showed that all 9 species of the endophytic bacterial isolates had the potential to produce antibacterial compounds against Escherichia coli and Staphylococcus aureus (Fatiqin, 2009). One characteristic of antagonistic microbes is that their growth rate is faster than pathogens and/or produce antibiotic compounds that can inhibit the growth of pathogens (Shehata, 2008).
Table (2):
Endophytic bacterial isolate ability to inhibit growth of Fusarium sp. and Colletotrichum sp.
| Isolates | Inhibition zone in cm | |
|---|---|---|
| Fusarium sp. | Colletotrichum sp. | |
| A1 | 0,5 | 0,4 |
| A2 | 0 | 0,9 |
| A3 | 0,2 | 0,8 |
| A4 | 0 | 0,5 |
| A5 | 0 | 0 |
| A6 | 0 | 0 |
| A7 | 0 | 0,6 |
| A8 | 0 | 0,2 |
| D1 | 0 | 0,6 |
| D2 | 0 | 0 |
| D3 | 0,6 | 0 |
| D4 | 0 | 1,4 |
| D5 | 0 | 1,4 |
| D6 | 0 | 1,0 |
| D7 | 0 | 0 |
| D8 | 0 | 1,2 |
The size of the inhibition zone produced is highly dependent on the type and stability of the antifungal metabolites produced by different bacterial species. This in vitro inhibition ability may be affected by the type, solubility, and stability of the antimicrobial compounds produced by endophytic bacteria. The stronger inhibition ability against pathogenic fungi is expected to be more effective in promoting plant growth by inhibiting pathogen attack plants.
Ability of Endophytic Bacteria of Blackboard Tree in Solubilizing Phosphate
The ability of endophytic bacteria in solubilizing phosphate may be due to their ability to produce phosphatase or extracelluler organic acids. The organic acid may be citric acid, glutamate, succinate, lactate, oxalate, glyco-oxalate, malate, fumarate, tartaric or alpha-ketobutyric acid (Maryanti, 2006), formic, acetic, propionic, lactonic, glycolic, and succinate acids capable of forming chelates with cations such as Al and Fe (Rao, 1994).
Six isolates A1, A3, A4, D1, D3, and D6 demonstrated to have ability in solubilizing phosphate showed by their PSI. A3 was observed to have higher PSI. The formation of clear zones around bacterial colony in Pikovskaya agar added with Ca3(PO4)2 (Figure 1) indicated that the isolates are capable of producing phosphatase or extracellular organic acids binding to Ca ions in the medium, liberating H2PO4 ions to form a clearer colored area compared to areas with bound P (George et al., 2002).
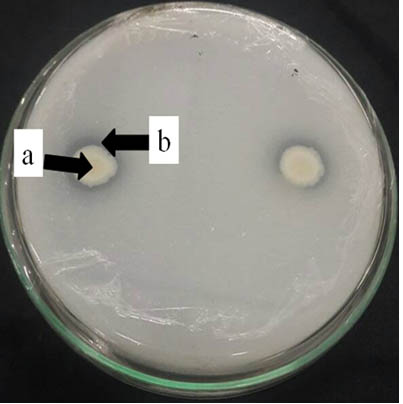
Fig. 1. Endophytic bacterial ability to solubilise phosphate shown by (a) clear zone around (b) bacterial colonies on Pikovskaya agar after 7 days incuba-tion at ambient temperature
Many studies were conducted to isolate endophytic bacteria from plants but blackboard. For examples 373 endophytic isolates was isolated from soybean plants, in which 49% were able to solubilize phosphate (Long et al., 2008). Similar study reported that of 77 endophytic bacteria isolates from Solanum nigrum (leunca), only six isolates were able to solubilize the inorganic phosphate (Rao, 1994).
Bacterial Growth and Production of IAA
A1 was observed to reach its maximum growth 20 hours after incubation. On the other hand, A3 was shown to reach its maximum growth earlier withn 12 hours after incubation before entering stationary phase (Figure 2).
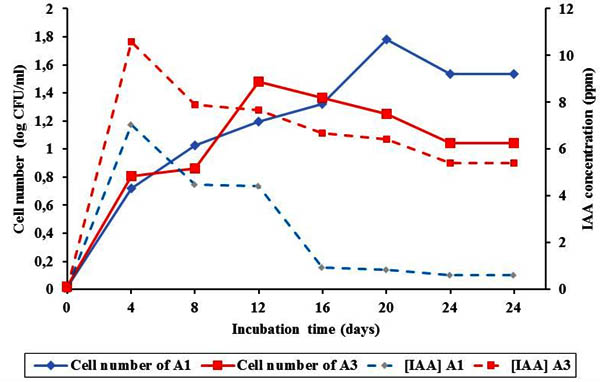
Fig. 2. Bacterial growth and IAA pro-duction of A1 and A3 for 24 hours
The process of elongation, cleavage and cell differentiation contribute by hormon such as IAA is better to support plant growth. In this study, IAA from endophytic isolates was measured using spectrophotometric method. IAA produced by A1 and A3 was varied. It was shown that both A1 and A3 produced high IAA in the first 4 hours and started to decrease there after. A1 and A3 might produce IAA with concentration of 7.041 ppm and 10.579 ppm respectively 4 hours after incubation. Decrement of IAA production together with increment of bacterial colonies indicated that some nutrients in the medium decreased during the period of IAA production (Lestari et al., 2007). Simultaneously, the bacteria mightreuse IAA due to nutrition deplition (Lay, 1994). Many studies were reported to isolate hormon producing endophyte bacteria. For example endophytic bacteria from corn roots produce plant growth regulator (Retnowati et al.,2011). Other study also reported that pepper plant fresh weight can be improved by soaking their roots in several endophytic bacterial isolates from pepper plant (Harni et al 2011). Similarly, endophytic bacteria from potato (Suriaman, 2010) and sweet potato (Hidayatullah, 2012) plants can produce IAA hormone at varied concentration to promote plant growth.
Assay on Application of Potential Endophytic Bacterial Isolatesin Chili
Crop production is more accurately expressed by measuring the dry weight than fresh weight. This is due to that fresh weight can strongly be affected by moisture conditions (Sitompul et al., 1995). Furthermore, the increment of fresh weight is influenced by the availability of sufficient and balanced nutrients due to increased cell division activity (Goldsworth et al., 1986). Dry weight results however are a balanced measurement between photosynthesis and respiration (Gardner et al., 1991).
Except that of A1F, other treatments of endophytic bacteria to chili increased fresh and dry weight of the plant. However, it varied among treatments with highershown by A3C (Figure 3). This might be due to differences in nutrient availability and micoorganism in the soil. Disease severity caused by pathogenic fungi might also contributed to fresh and dry weight. Compared to that of Colletotrichum sp. infestation, Fusarium sp. infestation might affect more to fresh weight that in turn affected dry weight. This fungus might decrease water intake by cloging xylem.
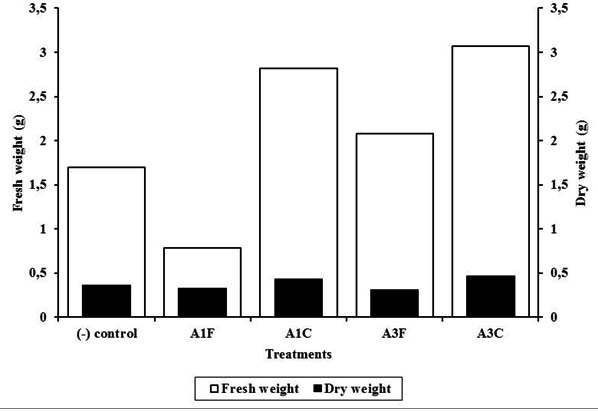
Fig. 3. Fresh and dry weight of infected chili plant treated with endophytic bacteria of blackboard tree
It was reported that many endophytic bacteria from different plants were able to solubilise phosphate to make it available for plants. Application of phosphate solubilizing bacteria increased paddy rice production by 5.4% (Thakuria et al., 2004). Moreover phosphate solubilizing bacteria applied in soil increased the fresh weight of red chili (Yanti et al., 2009). Another study showed similar result in which endophytic bacteria increased fresh weight and length of tomato roots (Munif et al, 2013). Pepper root and plant weight was increased by soaking the roots of pepper seedlings in the endophytic bacterial solution isolated from pepper plants (Harni, 2011). Fresh and dry weight of plant is the total weight of the plant showing the results of its metabolic activity. The availability of nutrients during plant growth leads to actively running photosynthesis (Sarief, 1986).
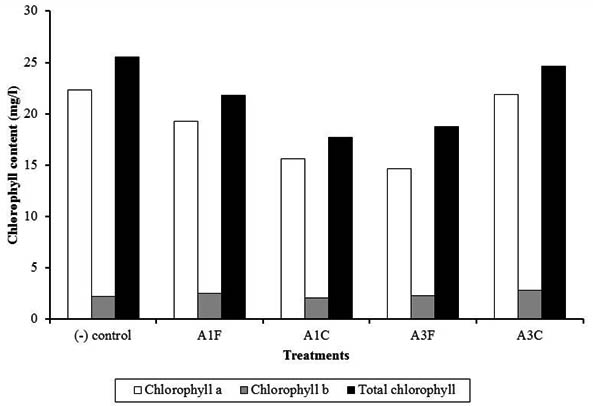
Fig. 4. Chlorophyll content of chili plants treated with endophytic bacteria of blackboard tree
A1F and A3C were observed to have highcontent of chlorophyll a and b, however they were lower to that of (-) control. Colletotrichum might cause leaf necrotics (antrachnose). In A3C, endophytic bacteria A3 probably decreased disease severity. Previous data showed that A3 was more capable to inhibit Colletotrichum rather than A1. Early symptoms of Fusarium wilt are pale leaf midrib particularly on apical regions, followed by epinasty on older leaves due to the of leaf petiole that causes the plants to completely wilt. Fusarium infection on young plants may cause immediate death due to the damages at stem base, while older plants could survive and but with very low production and small fruits (Semagun, 2000).
All treatments weredemontrated to increase plant height compared with (-) control (Figure 5). This suggested that application of endophytic bacteria were to promote plant growth. Plant height increment on treated plants might be due to IAA effect and phosphate solubilization by applied endophytic bacteria. Endophytic bacteria may increase plant growth by increasing the availability of plant nutrients such as nitrogen, phosphate, and other minerals, and stimulate plant growth by producing growth hormones such as ethylene, auxin, and cytokines (Bacon, 2007). Other results also reported that some endophytic bacteria can associate and promote chili plant growth (Sundaramoorthy et al., 2012).
Fig. 5. Plant height of chili plants treated with endophytic bacteria of blackboard tree
Number of leaves was varied between each treatment during observation. The highest average number of leaves increment was obtained from A3F treatment (Figure 6). Other treatments had lower average number of leaves increment compared to negative control. This indicates that A3F treatment is the best compared to other treatments.
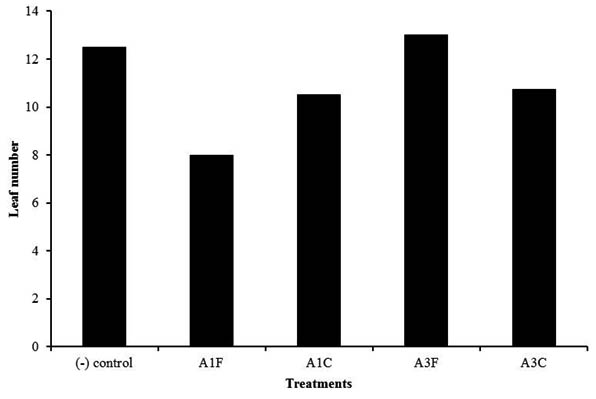
Fig. 6. Leaf number of chili plants treated with endophytic bacteria of blackboard tree
Application of endophytic bacteria was able to increase nutrient uptake of N, P and K. This increment was in line with the production of dry matter in plants. Nutrient absorption in plants is affected by nutrient content in the plant and the production of dry matter. Application of endophytic bacteria can promote the dry materials in plants (Gusmaini et al., 2013).
Bacterial cell number was counted before and 30 days after application using total plate count method on PCA and Pikovskaya medium. increased in all treatments, showed both on PCA and Pikovskaya agar(Table 3). Bacterial growth of phosphate solubilizing bacteria in this study was considerably low compared to that of Marista et al. (2013).
Table (3):
Bacterial count before and after application
| Code Isolates | PCA Medium | Pikovskaya Agar Medium | ||
|---|---|---|---|---|
| Number of cells before application (CFU/g) | Number of cells after application (CFU/g) | Number of cells before application (CFU/g) | Number of cells after application (CFU/g) | |
| A1F | 1,5 x 109 | 2,3x 1013 | 2,1 x 105 | 0,4 x 1013 |
| A1C | 1,3x 109 | 1,9 x 1013 | 1,8 x 105 | 0,8 x 1013 |
| A3F | 1,6 x 109 | 2,5x 1013 | 1,3 x 105 | 0,3 x 1013 |
| A3C | 1,9x 109 | 2,8 x 1013 | 2,8x 105 | 1,1 x 1013 |
Identification of Endophytic Bacterial Isolates Based on 16S rRNA gene
Both selected isolates, A1 and A3, were identified using 16S rRNA gene. Identification showed that A1 was relatively closed to Carnobacterium gallinarum strain DSM4847 with 84% gene similarity, while A3 was relatively closed Carnobacterium maltaromaticum strain JCM 1154 with 78% gene similarity. So far, these two bacteria have never been reported as IAA-producing bacteria nor phosphate solubilizing bacteria. Both species are gram-positive, lactic acid, non-spore forming bacteria. Previous study reported that Carnobacterium isolated from the atropic salmon fish intestinal was capable of controlling the growth of Vibrio anguillarum (Verschuere et al., 2000).
Sixteen isolates of endophytic bacteria were found from leaves and roots of blackboard tree (Alstonia scholaris). Two potential isolates, A1 and A3, showed to inhibit both Fusarium sp. and Colletotrichum sp. A3 was observed to have relatively high Phosphate Solubilizing Index and in producing IAA. It also showed that in general A3 promoted more chili performance, even with infestation of fungal pathogen. Identification using 16S rRNA gene sequence demonstrated that A1 was relatively closed to Carnobacterium gallinarum strain DSM4847 with 84% gene similarity, while A3 was relatively closed Carnobacterium maltaromaticum strain JCM 1154 with 78% gene similarity.
ACKNOWLEDGMENTS
We would like to thank to Universitas Sumatera Utara for partially supporting this research.
- Amilia E. Joy B. Sunardi., Residu Pestisida pada Tanaman Hortikultura (Studi Kasus di Desa Cihanjung Rahayu Kecamatan Parongpong Kabupaten Bandug Barat). Jurnal Agrikultura, 2016, 27(1), 23-29
- Bacon C.W. Hinton D.M., Bacterial Endophytes: The Endophytic Nische, its Occupants, and its Utility. In: Gnanamanickam SS. Gnanamanickam (ed.). Plant Associated Bacteria, Berlin, 2007, 155–194.
- Dewi T.K. Jodi S. Aguatiyani D., Isolasi dan Uji Aktivitas Bakteri Penghasil Hormon Tumbuh IAA (Indole- 3-Acetic Acid) dan Bakteri Perombak Protein dari Tanah Pertanian Tual, Maluku Tenggara.Pros Sem Nas Masy Biodiv Indon, 2016, 2 (2), 271-276.
- Dey R. Pal K.K. Bhatt D.M. Chauhan S.M., Growth Promotion and Yield Enhancement of Peanut by (Arachis hipogea L) Application of Gowth Promotion Rhizobacteria. Miicrobiolres, 2004, 159, 371-394.
- Eliza. Munif A. Djatnika I. Widodo., Karakter Fisiologis dan Peranan Antibiosis Bakteri Perakaran Graminae Terhadap Fusarium dan Pemacu Pertumbuhan Tanaman Pisang. J Hort. 2007, 17, 150-160.
- Fatiqin A., Isolasi dan identifikasi bakteri endofit dari daun dan kulit pulai (Alstonia scholaris) sebagai penghasil senyawa antibakteri terhadap bakteri Escherichia coli dan Staphylococcus aureus. Undergraduate thesis, Universitas Islam Negeri Maulana Malik Ibrahim, 2009.
- Gardner F.P. Pearce, R.B. Mitchell R.L., Fisiologi Tanaman Budidaya (Diterjemahkan oleh: Herawati Susilo), Universitas Indonesia Press. Jakarta, 1991.
- George T.S. Gregory P.J. Wood M. Read D. Buresh R.J., Phosphatase activity and organic acids in the rhizosphere of potential agroforestry species and maize. Soil Biol. Biochem. 2002, 34, 1487-1494
- Goldsworthy R.P. Fisher N.M., X Fisiologi tanaman budidaya tropik. Universitas Gadjah Mada, Yogyakarta, 1986.
- Gusmaini A.S.A. Munif A. Sopandi D. Bermawi N., Potensi Bakteri Endofit dalam Upaya Meningkatkan Pertumbuhan, Produksi dan Kandungan Andrografolid pada Tanaman Sambiloto. J Littri, 2013, 19(4), 167 – 177.
- Harni R. Ibrahim M.S.D., Potensi Bakteri Endofit Menginduksi Ketahanan Tanaman Lada terhadap Infeksi Meloidogyne incognita. Balai Penelitian Tanaman Rempah dan Aneka Tanaman Industri. J Littri, 2011,17 , 118 – 123.
- Hidayatullah F. Rahayu S.R. Lisdiana L., Produksi Hormon IAA Oleh Bakteri Endofit dari Akar Tanaman Ubi Jalar (Ipomoea batatas) dalam Media Limbah Cair Tahu. Lentera Bio, 2012, 2252-3979.
- Lay., Analisis Mikroba Di Laboratorium, Rajawali pers, Jakarta. 1994.
- Lestari P.L. Susilowati D.N. Riyanti E.I., Pengaruh Hormon Asam Indol Asetat yang Dihasilkan Azospirillum sp. terhadap Perkembangan Akar Padi. Jurnal Agro Biogen. 2007,3 (2): 66-72.
- Long H.H. Schmidt D.D. Baldwin I.T., Native Bacterial Endophytes Promote Host Growth in a Species-Specific Manner; Phytohormone Manipulations Do Not Result in Common Growth Responses, PLos one. 2008, 3(7), e2702.
- Marchesi J.R. Sato T. Weighmant A.J. Martin T.A. Fry J.C. Hiom S.J. Wade W.g., Design and Evaluation of Usefull Bacterium Specific PCR Primer that Amplify Genes Coding for Bacterial 16S rRNA. Applied Environmental Microbiology. 1998, 64: 795-799.
- Marista E. Khotimah S. Linda R., Bakteri Pelarut Fosfat hasil Isolasi dari Tiga jenis Tanah Rizosfer Tanaman Pisang Nipah (Musa paradisiaca var. nipah) di Kota Singkawang. Jurnal Protobiont, 2013, 2(2): 9-20.
- Maryanti D. Isolasi dan uji kemampuan bakteri pelarut fosfat dari rhizosfir tanaman pangan dan semak. Skripsi. Fakultas Pertanian Universitas Andalas, Padang, 2006.
- Munif A. Wiyono S. Suwarno., Isolasi Bakteri Endofit Asal Padi Gogo dan Potensinya sebagai Agens Biokontrol dan Pemacu Pertumbuhan. J Fitopatol Indones. 2012. 8(3), 57-64.
- Nusyirwani N. Kathy C., Isolasi dan karakterisasi bakteri Hidrokarbonoklasik dan Perairan Dumai dengan Sekuen 16s rDNA. Jurnal Ilmu Kelautan. 2007, 12(1), 12-17.
- Oktavia N. Pujiyanto S., Isolasi dan Uji Antagonisme Bakteri Endofit Tapak Dara (Catharanthus Roseus, L.) terhadap Bakteri Escherichia coli dan Staphylococcus aureus. Berkala Bioteknologi. 2018, 1(1).
- Pamungkas. Hakim L. Prastyono. Diro E.P., Data Base Jenis Jenis Prioritas Untuk Konservasi Genetik dan Pemuliaan. Departemen Kehutanan. Badan Litbang Kehutanan, Pusat Penelitian dan Pengembangan Hutan Tanaman, 2006.
- Pratyush K. Misra C.S. James J. Lipin D.M.S. Veettil A.K.T. Thankamani V., Ethnobotanical and Pharmacological Study of Alstonia (Apocynaceae). J. Pharm. Sci. Res. 2011, 3(8): 1394-1403.
- Radu S. Kqueen C.Y., Preliminary Screening of Endophytic Fungi from Medical Plants in Malaysia For Antimicrobial Antitumor Activity. Malaysian Journal of Medical Science, 2002, 9 (2): 23-33.
- Ramamoorthy V. Raguchander T. Samiyappan R., Enhancing resistance of tomato and hot pepper to Pythium diseases by seed treatment with fluorescent pseudomonads. Eur. J. Plant Pathol. 2002, 108(5), 429–441.
- Rao N.S., Mikroorganisme Tanah dan Pertumbuhan Tanaman. Edisi Kedua. UI-Press. Jakarta,1994.
- Retnowati Y. Wirnangi D. Putri S.H.E., Potensi Penghasil Hormon IAA oleh Mikroba Endofit Akar Tanaman Jagung (Zea mays). Universitas Negeri Gorontalo, 2011.
- Saraswati R.E. Santosa.Yuniarti E., Organisme Perombak Bahan Organik. Balai Besar Penelitian dan Pengembangan Sumberdaya Lahan Pertanian. Bogor, 2004.
- Sarief E.S., Kesuburan dan pemupukan tanah pertanian. Pustaka Buana, Badung, 1986.
- Semangun H., Pengantar Ilmu Penyakit Tumbuhan. Gadjah Mada University Press, Yogyakarta, 2000.
- Shehata F. Borollosy A.M., Induction of Resistance Against Zuccini Yellow Mosaic Potyvirus and Growth Enhancement of Squash Plants Using Some Plant Growth Promoting Rhizobacteria. Austr. J. Basic and App. Sci. 2008, 2, 174-182.
- Sitompul S.M. Guritno B., Analisa Pertumbuhan Tanaman, Gadjah Mada University Press. Yogyakarta,1995.
- Sundaramoorthy S. Raguchander T. Ragupathi, N. Samiyappan R., Combinatorial Effect of Endophytic and Growth Rhizobacteria Against wilt Disease of Capsicum annum L. caused by Fusarium solani. Biol Control, 2012; 60(1), 59-67.
- Supardi I. lingkungan hidup dan kelestariannya. Penerbit Alumni. Bandung, 1994.
- Suriaman E., Potensi Bakteri Endofit dari Akar Tanaman Kentang (Solanum tuberosum) dalam Memfiksasi N2di Udara dan Menghasilkan Hormon IAA secara In Vitro. Universitas Islam Negeri Maulana Malik Ibrahim, Malang, 2010.
- Tan R.X. Zou W.X., Endophytes: A Rich Source of Functional Metabolites. Nat. Prod. Rep. 2001, 18, 448-459.
- Thakuria D. Talukdar N.C. Goswami C. Hazarika S. Boro R.C., Characterization and Screening of Bacteria from Rhizosphere of Rice Grown in Acidic Soils of Assam. Current Science. 2004, 86, 978-985.
- Verschuere L. Rombout G. Sorgeloos P. Verstraete. W., Probiotics bacteria as biocontrol agents in aquaculture, App. Environ. Microbiol, 2000, 64: 655-671.
- Yanti Y. Gustian H. Rahma., Aplikasi Agen Hayati Pseudomonas fluorescens Sebagai Penginduksi Ketahanan Untuk Meningkatkan Produksi Tanaman Cabai Terhadap Penyakit Virus Kuning di Kecamatan Kuranji Kotamadya Padang, Warta Pengabdian Andalas, 2009, 15(22), 48-54.
- Zinniel D.K. Lambrecht P. Haris N.B. Feng Z. Kuezmarski D. Higley P. Ishimaru C.A. Arunakumari A. Barletta R.G. Vidaver A.K., Isolation and Characterization of Endophytic Colonizing Bacteria from Agronomic Crops and Prairie Plants. Applied and Enviromental Microbiology. 2002, 68 (5), 2198-2208.
© The Author(s) 2018. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.